
|
|
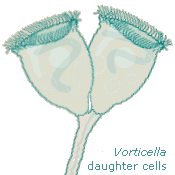
Sessile peritrichs are ciliates that achieve a more settled lifestyle than most other protists by attaching themselves to something more stable than they are. Spirally arranged rows of cilia encircling their disk-shaped front ends create powerful little vortices in the water and draw food particles to their mouths. Many of the sessile ("seated") peritrichs, like Vorticella, grow stalks (Leeuwenhook called them "tails") that anchor them in place. Telotrochs are what peritrichs turn into when they need to move. |
 |
|
Half the population needs to move once reproduction is accomplished—two daughter cells can share the end of a stalk for just so long, and then one has to leave. When there's too little food or some other undesirable situation nearby, adult peritrichs can make themselves into telotrochs and swim away. Then they grow stalks and settle down again. Usually. | ||
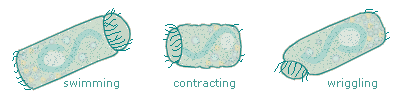
This process is fascinating, but it's also problematical for us because the telotrochs don't necessarily look like the sessile peritrichs they once were. Several common species of Vorticella that look like those daughter cells elongate themselves considerably when they're traveling; as such they lose their familiar, identifiable bell shape. Further, they seem to contract themselves even more often than when they're sessile, and sometimes they stop and wriggle, so there's not even one cylindrical outline to watch for. |
 |
|
When telotrochs contract and wriggle, they cause interesting things to happen to their pellicles. Contraction affects the rear end more than the front; a good, strong contraction puts accordion pleats into the posterior half of the telotroch. |
 |
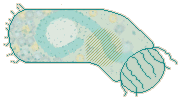 |
When they wriggle energetically, telotrochs bend their bodies to the degree that faint little striations appear on the side toward which they're bending. This implies that the front of the telotroch is not so elastic as its other end. |
|
It's understandable that, up front, teletrochs are not quite the shapeshifters that they are farther back. The front of the telotroch, when it used to be the rear of the sessile zooid, needed only to contain cytoplasm and connect the contractile stalk to the contractile peristomal area; that part of the pellicle itself did little contracting or expanding. Those wonderful accordian pleats in the pellicle of the telotroch's posterior half have just the opposite history. |
||
 |
When a sessile Vorticella's stalk contracts, its peristome changes shape markedly. The "lips" from which the cilia grow contract when the stalk does, pulling the cilia inside the "bell." Instead of being aligned back to front, as they are normally, the cilia are turned 90° and point toward the center of the circle defined by the "lips"; depending on how long the cilia are and how strongly the organism contracts, their ends touch or even overlap. Meanwhile, the pellicle that was just below the peristome when the Vorticella was extended contracts and virtually closes, like the draw-string closure of a laundry bag. When this region becomes the rear end of a telotroch, it retains its elasticity and can easily fold into those accordian pleats.
|
Aside from my misinterpretations of what I'd been seeing and a little bit of anthropomorphism, I had some reason to delay my conclusion about telotrochs. In books that mention and picture telotrochs at all, there's no sign of the cilia that I've seen on their rear ends. Jahn depicts the formation of a telotroch in these five stages (Figure 444 in How to Know the Protozoa): |
 |
Kudo's depiction (in Protozoology) includes two earlier stages and the magic words "in vitro." So these are "test-tube telotrochs" (sorry; I couldn't resist), and different conditions may produce a somewhat different sequence. Of course, I didn't notice that qualifying phrase in Kudo's caption the first time I looked at the drawing, but once I did, it became a source of consolation. The telotrochs I'd been seeing were variants, that's all. And different species of Vorticella—as well as different genera and families of peritrichs—would probably include other variations. |
Another conclusion I still have difficulty affirming is that free-swimming peritrichs don't use peristomal cilia for locomotion. The organisms to the right may or may not be telotrochs. They'd been so numerous on many of my slides that, before I learned to use methyl cellulose to slow down swimming micro-critters, I thought they were a new kind of protozoon. Then I recognized them as Vorticella without stalks, read what I could find about them, and began my acquaintance with telotrochs. |
 |
|
I learned how peritrichs leave their stalks behind to swim to a better location, how they'd change their appearance, how their peristome wasn't apparent anymore, how they grew cilia on their rear ends, and how they used these new cilia to get around. Trouble was, the organisms in this drawing weren't explained by what I'd been reading. |
||
Then, for two months last summer, the water I collected contained large populations of peritrichs. And if one perseveres ... yes, I found a Vorticella changing unmistakably into a telotroch! As I watched, the little stalk-nub smoothed over, the cilia around it grew, and the peristomal cilia ceased motion; the new cilia clearly began to move the organism. |
 |
Still, the telotroch above became detached from its stalk before its new ring of cilia became operative. Although I had not seen it swimming before I watched its new cilia grow, any traveling it did before I came upon it had to have been done by its peristomal cilia. I've since seen stalkless Vorticella and Epistylis swim by using just their peristomes, and I've even found a Vorticella with none but peristomal cilia towing its stalk behind it and, behind that, a small bit of the substrate to which it was still attached. The inevitable conclusions are (1) Vorticella do turn into textbook-variety telotrochs, and (2) with or without their stalks, if they don't have trochal (posterior) cilia they can get around quite well by using their peristomes.
|
The many peritrichs quickly taught me that my earlier identification of the "new kind of protozoon" as a stalkless Vorticella had been more comforting than accurate. No doubt some of them were Vorticella, but others could have been the colonial peritrichs Epistylis, Carchesium, or Zoothamnium, or the solitary Apiosoma or Campanella. (2)
|
 |
 |
 |
 |
(Peritrichs' contractile stalks don't have myonemes inside them anymore. As the elastic fibers are made of the protein spasmin, not acto-myosin, the stalks contain spasmonemes.(3)) |
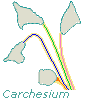
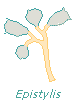
Vorticella are independent, each zooid having a contractile stalk entirely to itself. The individual organisms are nevertheless gregarious, usually found in the company of others. Carchesium are colonial creatures that share stalks. Within the shared stalk, however, each Carchesium has its own elastic filament (the more intense colors, above) that enables it to contract (see the bottom zooid) without causing the other members of the colony to contract with it. Zoothamnium are colonial and share stalks, too, but the elastic filament of each zooid is connected to those of the others; when contraction occurs, the elastic fibers pull all the organisms toward the site where the stalk is attached to the substrate (all zooids shown are extended). Epistylis are colonial, with stalks joined to those of other members of the colony. But since their stalks contain no elastic fibers, contraction affects only the individual zooids (in the drawing, half are contracted).
|
From what I've been able to find written about them (4), I removed Carchesium from suspicion; they don't become regular teletrochs. |
 |
If this pair I watched were Carchesium, both daughters grow stalks that separate them within the colony. |
||
If not Carchesium, they were probably Zoothamnium because of how common the latter are and because I didn't find members of other genera that were colonial with at least somewhat contractile stalks. (When I was watching them reproduce, they were under a coverslip; otherwise I could have poked them gently and let their response identify their genus.) The likeliest suspects thus became Epistylis. (5) |
||||
Reading, I learned that Epistylis are not monomorphic as Carchesium are, that they do have a teletroch stage. Observing, I noticed things that began to draw my questions and tentative conclusions together. For one thing, unlike Vorticella, the free-swimming Epistylis I watched were fatter than sessile ones. |
 |
 |
These Epistylis (probably E. niagara), their relative sizes scrupulously shown, were drawn from a number of individuals. The sessile one (left) was 78 µm long, 35 µm at its widest diameter. The others are side- and from-the-top views of a free-swimming zooid that was 60 µm in diameter and 28 µm thick (peristome to scopula, where the stalk attaches); others ranged from 40 to 65 µm in diameter, and all were manic swimmers.
|
While I had not seen any Epistylis transform themselves into the telotroch form shown above, I had been seeing many peritrich swimmers that I thought were Vorticella; less ignorant about peritrichs, I began to find numerous Epistylis that weren't fat telotrochs but that lacked stalks, like this one. |
|||
 |
It had the identifying three turns of peristomal cilia, extra "lip" or "collar" just below the peristome, and long, curved cytopharynx or gullet, just as stalked Epistylis on the same slides had. Besides the presence or absence of a stalk, the chief difference between them was that the free-swimming zooids remained a bit contracted, slightly longer front to back than they were in diameter (125 x 95 µm). When these swimmers truly contracted, pulling in their peristomal cilia, they were only 100 µm long and fully 110 µm across. |
||
Here are the same stalks and zooids as are in the drawing of Epistylis' outline and stalk configuration above. |
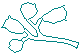 |
Notice how the zooids seem too large for the stalks. I thought I'd drawn them wrong, and I double-checked. |
||
The proportions are correct; while it's usually silly to make judgments about the way other living things look, in this case the anthropocentric "reflex" can be made to serve a purpose. Epistylis "bells" and stalks are, it seems, connected tenuously enough that the "bells" break free with relatively little provocation. |
||||
Thinking back on the peritrichs I had transferred from my water jars to slides, I realized that (1) looking at the colonies in the jars with my stereo microscope I saw few if any of the stalkless zooids; (2) transferring colonies of Carchesium and Zoothamnium to slides and observing them with my compound microscope, I found only a few of the stalkless and trochal-cilia-less organisms; and (3) doing the same to colonies of Epistylis resulted in hordes of stalkless zooids that used their peristomal cilia to move about.
|
In November, amid exposed rocks below the small (six feet high) dam on the slow-moving Lost River, I took water from two isolated pools in the streambed. The river was much lower than I've ever encountered it, and I had no idea of what life it was harboring. Green life: desmids, green hydras, crustaceans and rotifers with green bellies. At home, once the water had settled a bit, I used my stereo microscope and found a large, golden-green rotifer. |
 |
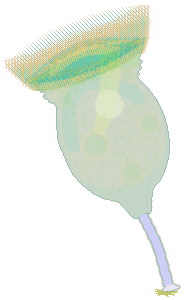 |
After I transferred it to a slide, I saw what you noticed right away: the 250-µm-long "rotifer" was a gigantic peritrich. Its body was 170 µm in diameter, and it had a 110-µm-long stalk of refractive, crystalline blue, at the end of which was a holdfast still attached to a piece of substrate. It was relatively motionless, contracting itself occasionally so that its peristome was only 110 µm in diameter, then extending its long peristomal cilia and ingesting whatever they brought into its cytopharynx. This was a beautiful Campanella umbellaria, a big "stentor" of a peritrich, so large that the transverse marks on its pellicle were visible ("lines" 3 µm apart). I could see how the cytoplasm was denser at its posterior end, and how, on the right and left sides of its body, a similar darkening occurred where the illumination had to pass through more pellicle than it did nearer the middle. |
|
Even the spasmonemes showed as faint lines radiating from where the stalk attached, upward and toward the organism's sides. (So they are the means by which peritrichs contract themselves!) At the end of the cytopharynx food vacuoles formed, about 20 µm in diameter, and moved counterclockwise, becoming smaller as digestion proceeded. The contents of the food vacuoles contributed to the organism's overall color, but the green hue was established by numerous small zoochlorellae: the size of this Campanella allowed me to see clearly that they were not enclosed in food vacuoles but were circulating in the cytoplasm. The size of the organism also revealed the extra "lips" at the base of the peristome. I wasn't able to determine how many rows of peristomal cilia there were, but (again, because of how big this organism was) I could see that there were more than three. I could also see that extension and contraction did not affect the shape of this Campanella except from its "lips" forward, where the cilia changed from being almost mesmerizing in their graceful motion (peristome extended) to appearing very much like an unruly haystack when the peristome was not quite fully contracted and individual cilia were oriented in every possible direction.
|
||
The big Campanella had trochal cilia! They were barely visible, 6 µm long, growing along the border of the denser cytoplasm near its posterior end, 30 minutes after I'd pipetted the organism from my water jar onto the slide.
|
||
At 40 minutes, the cilia were 9-12 µm long; the stalk was still blue, but less vibrantly so, and it looked as if it were breaking into segments.
|
 |
|
 |
At 2 hrs 11 min (½ hr later), the trochal cilia were gone and a new stalk was growing: 22 µm long and 22 µm (the old stalk had been 15 µm) in diameter.
|
At 6 hrs, the stalk was 165 µm long and no longer attached. The Campanella had grown a new one that was 30 µm long and, like the previous one when it was short, that looked very much like a tail. Neither of these stalks had a holdfast. Both were mostly transparent, without any refractive color. This time (4½ hours, from when it had detached from its first stalk) the Campanella had traveled 750 µm.
|
Remembering that Epistylis teletrochs seem the result of the stalked zooid's having been squashed from front to rear and that Vorticella transform themselves into "cylinders" that look considerably longer and narrower than the zooids were while sessile, I was probably expecting the Campanella to alter its shape. To the right is how the telotroch typically looked. |
 |
|
If you look closely, you can see how the organism's squared-off or rectangular appearance is mostly an optical illusion: when the trochal cilia flick out, they create a "corner" that sustains the illusion, which is reinforced by the peristomal cilia's no longer being extended in a series of regular, graceful lines. But the difference in shape between the Campanella with a stalk and peristomal corona and how it looks when it has trochal cilia and no stalk is more apparent than real. |
||
 |
|
Here they are together: the Campanella immediately after it lost its stalk and the same critter several minutes later. Excluding peristomal cilia, both images are 59 pixels across. When I measured the images, the result was a surprise. |
||
Of course, there is an actual, small change in shape. In the rear the scopula is pulled "in" or forward, and in front the peristomal cilia are no longer fully extended. The partial contraction of the whole organism accounts for both these results and also for the thickening of the body just behind the peristome. I'm sure there's a relation between this contractedness and that of the swimming, stalkless Epistylis I'd been finding, but I don't know what it is beyond the fact that neither those swimmers nor this telotroch had closed off their anterior ends the way that Vorticella teletrochs typically do. |
||||
This Campanella telotroch exhibited three forms: a "proper" telotroch form in which all the peristomal cilia were drawn into the anterior part of its body, a sessile-like form in which those cilia were fully extended, and a partly contracted form in which those cilia were, at least visually, in a state of disarray. Watching this Campanella, I couldn't suppress my judgment that it didn't know what it was doing —or, to use less subjective language, that it behaved as if neither the sessile nor the telotroch form was a satisfactory response to its environmental situation. When it had no stalk and was traveling (in its most compact form, with the trochal cilia moving it), the organism would nevertheless extend its peristomal cilia either part way or all the way every few moments; once the cilia were out and working, the Campanella was feeding. Particles were arriving in its cytopharynx and being packaged into food vacuoles even when its peristome was disarranged. Was the organism accumulating information about the availability of food in its current location? —certainly. Was it acting on the information? —who knows?
|
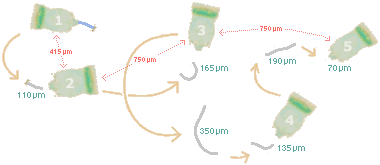 |
The Campanella had also dropped 13% of the volume it had when it arrived on the slide, and 23% of the volume it had 17 hours earlier. Its food supply had run out, and there probably wasn't much oxygen left (most of the slide's other occupants had succumbed to anoxia). But it continued to use its peristomal cilia and to create food vacuoles. In contrast to the day before, the vacuoles were practically empty. Probably, these food vacuoles were pinching off when they reached a certain volume irrespective of their contents. The area around the Campanella was relatively free of excreta; in fact, the little clumps were most numerous around the organism's first two positions (on the "trail of the telotroch" above) and gradually, in an almost even distribution, grew fewer in number as it moved on. Considering this pattern, the diminution of the telotroch, and the way those studied Vorticella were made to sever their stalks electro-chemically, I wonder if there isn't a relation. Since digestion changes the pH of food vacuoles, the Campanella may have become a telotroch as a response to the the water in the immediate vicinity of its excreta. Reaching somewhat "better" water, it grew a stalk. Then it responded to the lack of food in the new location by growing trochal cilia and dropping the stalk, and then it responded to the still "better" water by growing a new stalk, and then to its "still hungry" condition with new trochal cilia, and then.... —No wonder the critter was losing weight! |
 |
It was still quite a beautiful "animalcule." It had provided particularly easy-to-see illustrations of telotroch morphology and behavior, some of which I've already been able to relate to subsequent sightings of smaller, less revealing telotrochs. Responding to the excellent observations it had afforded me, I gently rinsed both slide and coverslip into the water jar, where, I hope, the Campanella found something to hold fast to and many little somethings to eat. |
Notes |
Comments to the author Rosemarie Arbur welcome.
Please report any Web problems or
offer general comments to the
Micscape
Editor,
via the contact on current Micscape
Index.
Micscape is the on-line monthly magazine
of the Microscopy UK web
site at Microscopy-UK
WIDTH=1