
|
PHOENIX RISING Eclipse and resurrection of a diatom sanctuary. Michael J. Stringer (UK) and Frithjof A.S. Sterrenburg (The Netherlands) |
(Click on diatom images for larger images showing frustule detail.)
The Sanctuary
The diatom sanctuary discussed here is a tiny lagoon on Two Tree Island, which lies in the Thames Estuary between Leigh on Sea and Canvey Island in the county of Essex, UK (Figs. 1). The island is very small, is uninhabited by man but is home to thousands of wintering migratory birds that arrive from the North of Europe and Russia. In the Spring, when these depart, they are replaced with arrivals from the South, for breeding.
Figure 1
View Larger Map
Green arrow shows the coordinates 51°32'
21.97” North, 00° 38' 21.97" East.
Embedded map coding courtesy of Google Maps.
The habitat is essentially tidal marine, accessing hundreds of small lagoons, but there are also some small pockets of brackish water and two quite large freshwater lagoons. The diatom diversity is very large for such a tiny location.
(Michael Stringer’s story):
This paper focuses on just one of these lagoons from which I regularly collect, which I call The Car Park site. It is very small, only 20 x 20 metres with one gully to a creek and then the Thames. I would regularly collect samples from this and the other sites, every 14 days or so, in the manner of Dr. C.L. Odam who collected samples from the River Eden — which is a tributary of the Medway, almost opposite Two Tree Island. in Kent.
Sampling procedure
My collecting procedure has been developed over the years and, as they say, it works for me. It is very simple and can easily be adapted for your own use. To reach out into awkward places, I use a 16 foot telescopic carbon fibre fishing-rod with a little 2.5” square stainless steel scraper on the end. This is actually a very small cheese grater, £1 from The Factory Shop. A 2.5” sliver of mud is removed with this and is transferred to a little pot. I have no idea what the original use was of these little pots but they are perfect for diatom traps. The lid is transparent and allows in plenty of light. I use a collecting box with twelve compartments, which also contains some 50 ml syringes, brushes, notepad and pens; thermometer and hydrometer to take both air and sea temperatures, weather conditions and an hydrometer reading.
At home, the pots are tapped to settle the mud down into the base and also flatten the surface. The mud is then covered with one, or sometimes two, layers of gauze mesh, width about 150 – 200 microns, which is then moistened. The lids are secured in place and the pots are then placed in the daylight (Figs. 2). The gauze is regularly checked with a microscope for diatoms that have climbed out and photographs are taken. If the diatoms are very actively motile, I take an AVI video as well. The first diatoms to make an appearance are tiny “Naviculas”, Amphoras and Nitzschias, usually within 40 to 45 minutes. Larger species, Gyrosigma and the like, start to appear within 12 hours. The gauze is changed every 48 hours because worms, snails and other unwanted customers are then appearing, which make cleaning more difficult.
The samples are cleaned with H2O2 and dichromate, washed and strews are mounted with Zrax or Pleurax.

Figure 2
History
About two years ago, the diatom population began to wane and in 2010 it rapidly collapsed altogether. The picture (Figs. 3) was taken in February 2009 when, normally, the Spring Bloom would already have been kick started. There are no tell-tale signs along the margins, in particular, of any diatom colonies. Normally, this would be a deep-brown marginal layer extending into the lagoons by quite a few feet. At about the same time, the Royal Zoological Society mentioned on the BBC News that the Annual Eel Count of the river Thames was mysteriously down by 98%. I sent them an email informing them I had observed that the diatoms in the Estuary had also declined by the same amount but got no reply.

Figure 3
Now, another strange coincidence. I was talking to one of the local cockle fisherman and he told me that he has never known seasons as bad as this and that is going back for more than 70 years! He described the river as being “dead” and suggested that this was caused by the dredging of an enormous channel for giant ships for a deep water harbour at Shell Haven, a few miles further up the river. The Thames is about 11 metres deep and the channel to be created will be 16 metres deep. Work started in March 2010 and probably earlier than that.
It is only in recent years that the Thames has become clean and salmon have been seen running up the river. Before that, it was a sewer, foul stinking water. Raw sewage and industrial waste were simply piped into the river by all the communities along its banks. The water quality was awful and one can only imagine what the silt contained. Five metres of silt, at least one hundred metres wide and many miles long containing all sorts of pollutants, will be released over the following months. The main dredging will start in March 2011 and I will be watching the diatom populations around the island very carefully.
On the 14th November 2010, the weather and tides were favourable for a regular check, starting at The Car Park, where I was greeted by diatom blooms! (Figs. 4). What a pleasure after a lapse of almost two years and I managed to collect two samples. I moved around to another locality and the colonies were even more spectacular. This was just like the old times! I spent so much time there that the tide was coming in very quickly and I had to leave the other sites until another day. That wasn't to be until the 15th December, during the recent bitterly cold weather we have had in the UK. I had a chance to collect more samples and also visit other sites that had been barren for the past year and there was a faint bloom here, too.
Remarkably, all 12 sites I examined had diatoms re-established, despite temperatures being down to -8ºC. with a couple of feet of snow. In the interim period, there were no signs of diatoms, just the grey mud, which may have contained diatoms. The typical black bloom, complete with oil slick, had not been seen for many months. The good news had to be shared with Frithjof and samples and photographs were sent off.

Figure 4
(Frithjof Sterrenburg’s story)
A winter’s crop
To illustrate the recovery of the diatom crop, here are images of some members of the flora in the sample from “The Car Park Lagoon”, Two Tree Island, Essex, U.K., collected on 14th Nov. 2010. They are illustrated in ordinary brightfield, without any optical or digital trickery except for some adjustment of brightness and contrast. The illustrations therefore correspond to the “natural” aspect on visual observation. I find few things more tedious than the countless applications and removals of immersion oil when switching between scanning of the slide at low power and photomicrography at higher power, so all pictures were taken with a 50/1.00 water immersion for convenience. For most of the species encountered, this is quite sufficient.
The method of sampling (see above) favours the presence of actively motile raphid species, but occasional centrics (e.g. Actinoptychus undulatus Ehrenberg) and araphid pennates (e.g. Rhaphoneis amphiceros Ehrenberg) are also present. The total number of frequently encountered species in the resurrected flora is about 30, which corresponds to a good diversity for such an environment. In addition, numerous less frequent species are present, e.g. from the genus Fallacia, and many other small Naviculoid and Nitzschioid diatoms still awaiting identification.
The combination of Caloneis westii (W. Smith) Hendey, Parlibellus plicatus (Donkin) Cox, Gyrosigma balticum (Ehrenberg) Cleve and Pleurosigma angulatum (Quekett) W. Smith as the most abundant species is sufficient to immediately recognize the nature of the sample as “a typical saltmarsh as seen on the coasts of W. Europe”.
Caloneis westii (formerly also called C. formosa) (Figs. 5) is common in brackish littoral waters. Note that there are clear indications of nearly-resolved areolae in the striae, despite the limited NA of the objective used.
Parlibellus plicatus (formerly called Navicula plicata) (Figs. 6) is also widely encountered in the marine littoral. The valve is extremely vaulted and for a total survey, stacking of several images would be required. The view is on the girdle, showing the rows of fine foramina in the girdle bands. In an oblong field I have increased contrast to better show the areolae.
Pleurosigma strigosum W. Smith (Figs. 7) is not to be confused with P. angulatum: it’s immediately evident that the former shows no switch to perpendicular striation at the apices. In the inset I have greatly enlarged a portion of this image to illustrate an interesting optical phenomenon. In SEM, the outer layer of the sandwich valve is seen to be perforated by tiny slits, the inner layer by much larger round openings —the “pearls” in the usual LM image. The slits are here visible as narrow black “scratches”, the round openings as the surrounding much larger grey shadows. With very careful focusing, you can try Gyrosigma balticum for an easier experiment.
A muddy broth like this sample often yields a variety of Gyrosigma species, and half a dozen have so far been observed in the sample.
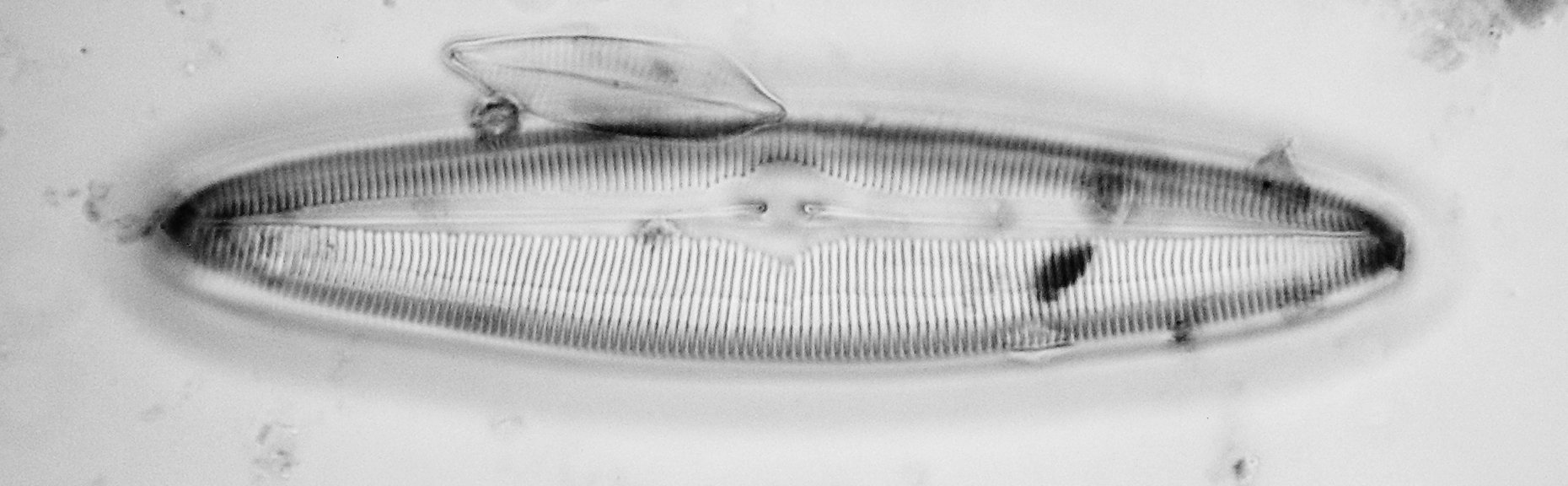
Figure 5 - Caloneis westii

Figure 6 - Parlibellus plicatus
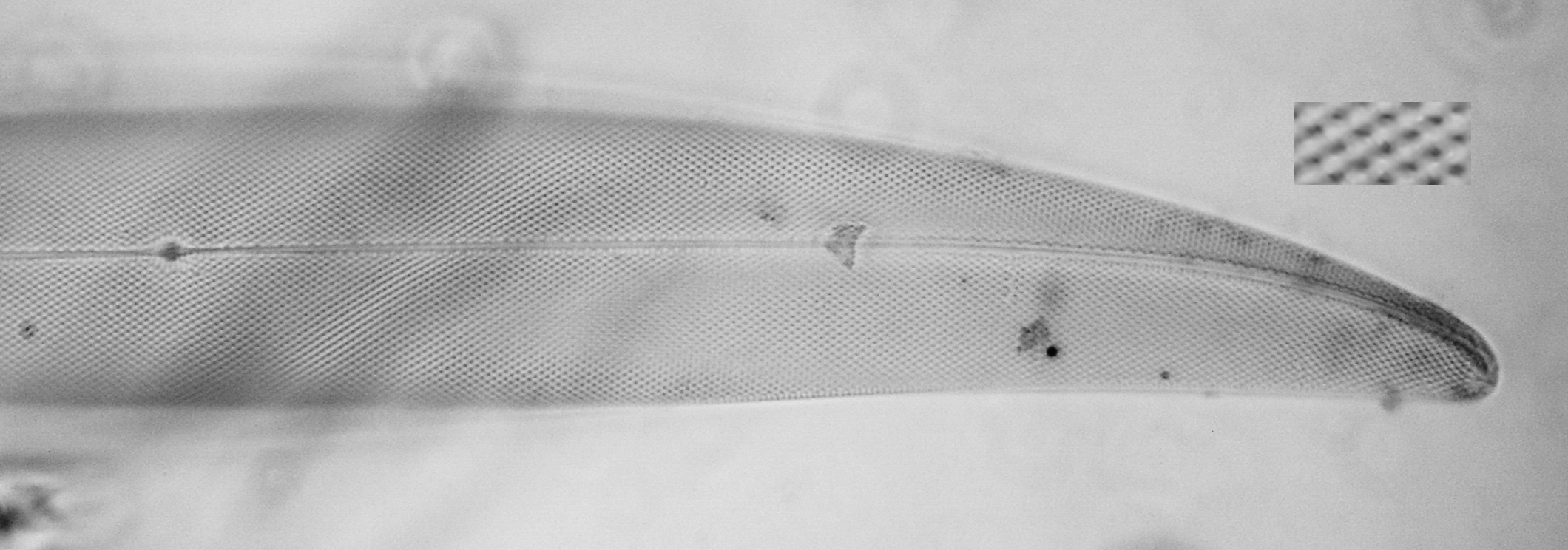
Figure 7 - Pleurosigma strigosum W. Smith
Gyrosigma littorale (W. Smith) Cleve (Figs. 8) is easily recognized by its very coarse longitudinal striation. I have regularly encountered it, but never in large populations.
Gyrosigma wansbeckii (Donkin) Cleve (Figs. 9) looks like a small Gyrosigma balticum but has much finer striation (ca 20 in 10 µ). It is frequently present in such habitats, but apparently often overlooked.
Figs. 10 shows a species resembling the previous, but differing in various features including even finer striation (ca. 25 in 10 µ), which most probably corresponds to Gyrosigma lorenzianum Grunow. It has rarely been mentioned, probably because of this similarity, but is not at all rare, also in this sample.
Gyrosigma scalprum (Gaillon) Sterrenburg (Figs. 11) had for some 150 years been a victim of the unholy confusion around “G. hippocampus” — which name, despite being invalid, had been used for various diatoms — but it is a clearly individual entity and I therefore formally defined it as such in 1992.
Petrodictyongemma (Ehrenberg) D.G. Mann (Figs. 12) is the current name for the classic “Surirella gemma” and like Diploneis didyma (Ehrenberg) Cleve (Figs. 13), it is a common mudflat dweller.
Scoliotropis latestriata (Brébisson ex Kützing) Cleve is not so frequently seen and is challenging because it is very strongly vaulted in both valve (Figs. 14) and girdle views (Figs. 15). The extreme blow-up of Figs. 16 is a girdle view showing the typical double rows of areolae with an apo 100x/1.4, which of course has an even smaller depth of field. If I find particularly clean specimens in the sample, they would be good candidates for stacking half a dozen or so shots at different focus settings.

Figure 8 - Gyrosigma littorale (W. Smith) Cleve

Figure 9 - Gyrosigma wansbeckii (Donkin) Cleve

Figure 10 - probably Gyrosigma lorenzianum Grunow
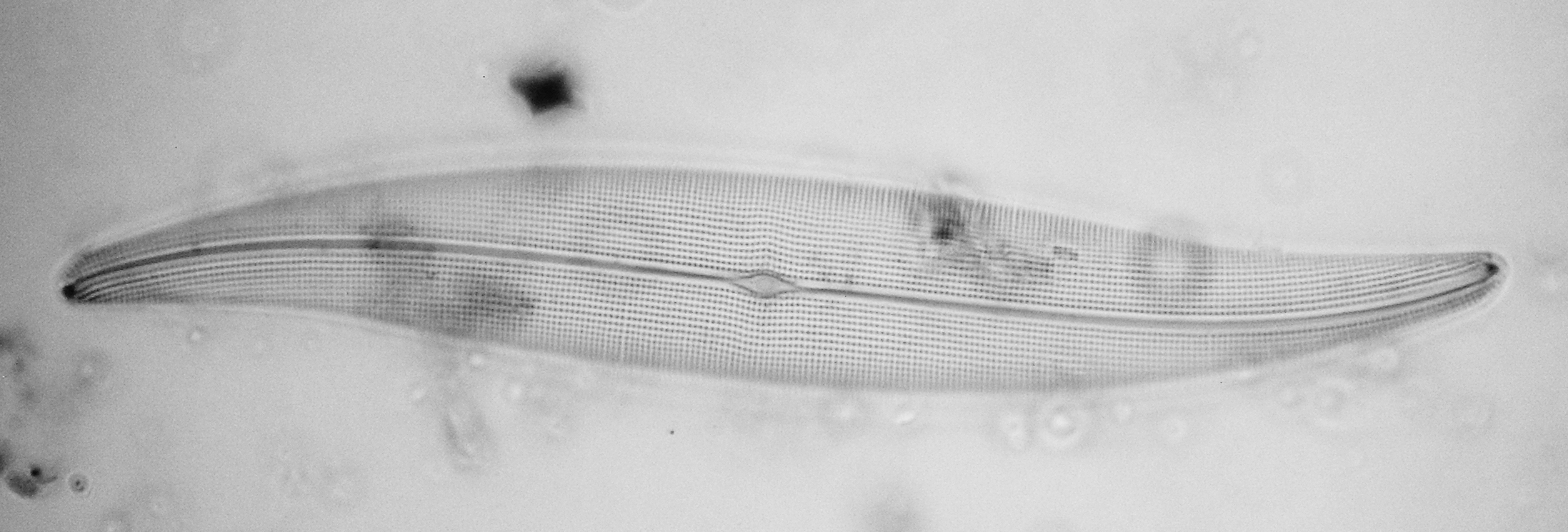
Figure 11 - Gyrosigma scalprum (Gaillon) Sterrenburg
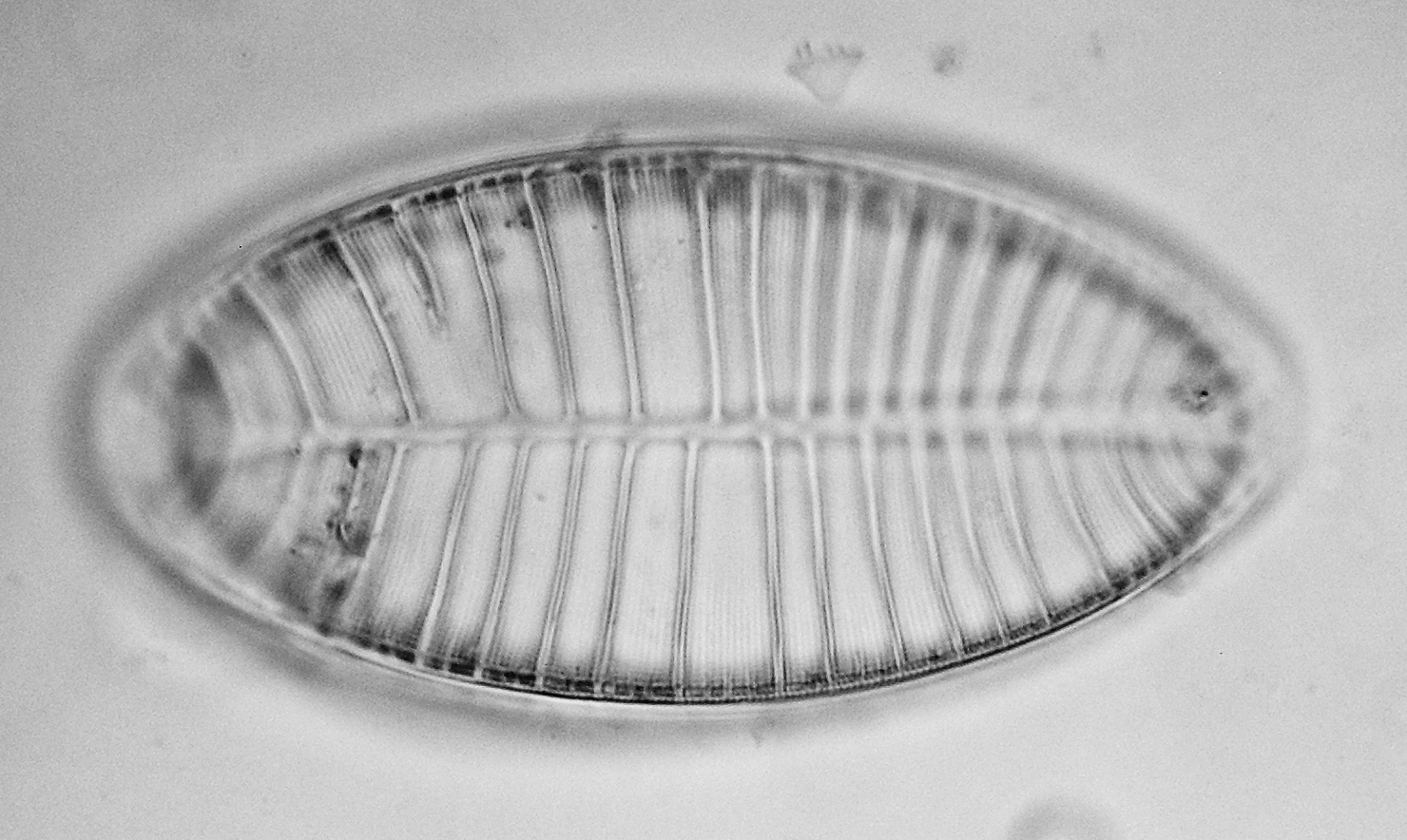
Figure 12 - Petrodictyongemma (Ehrenberg) D.G. Mann
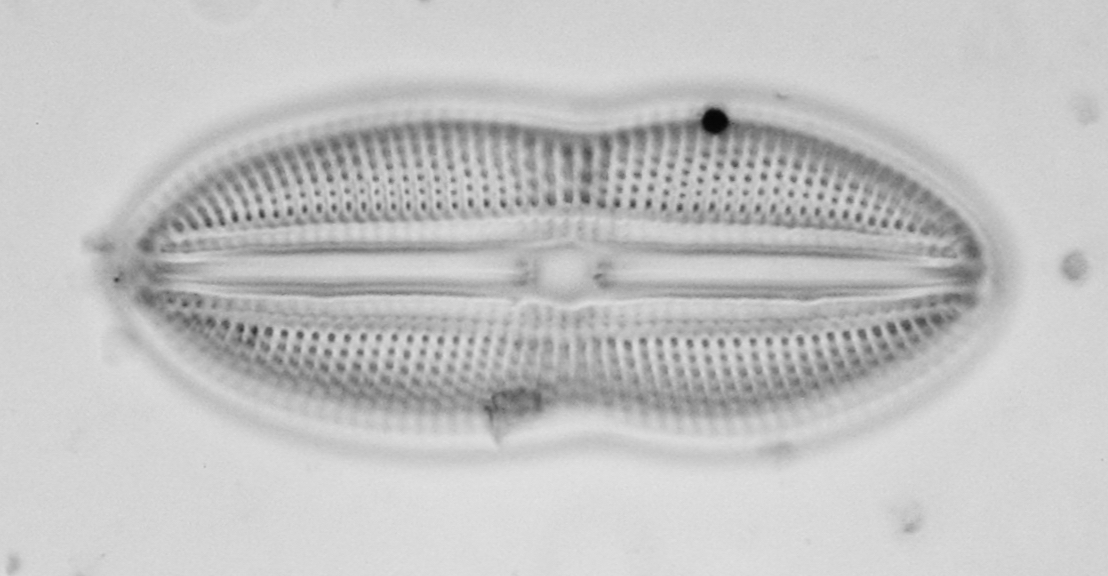
Figure 13 - Diploneis didyma (Ehrenberg) Cleve
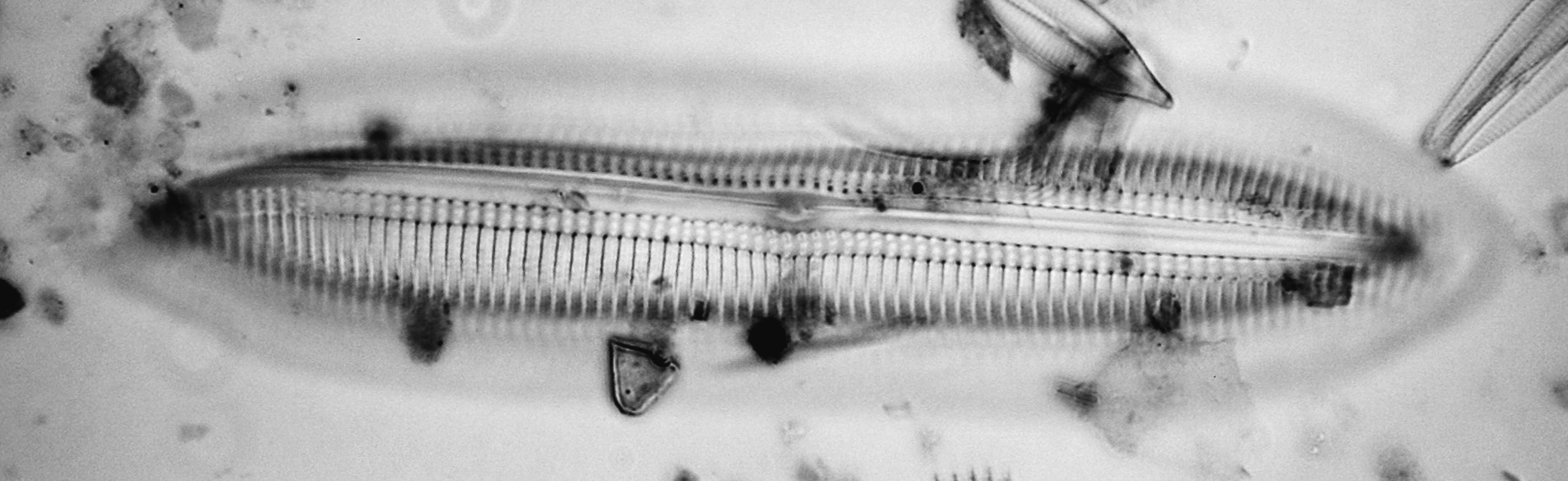
Figure 14 - Scoliotropis latestriata (Brébisson ex Kützing) Cleve
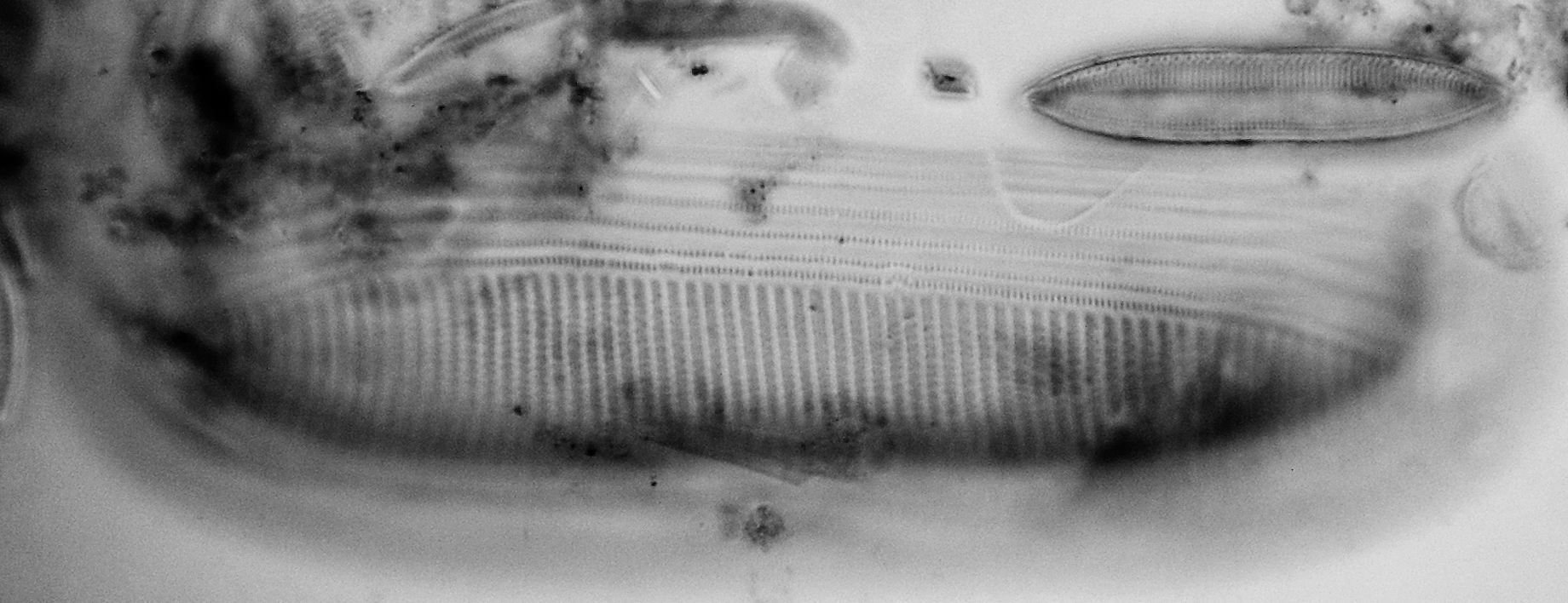
Figure 15 - girdle of Scoliotropis latestriata (Brébisson ex Kützing) Cleve

Figure 16 - magnified girdle detail of Scoliotropis latestriata (Brébisson ex Kützing) Cleve
Population dynamics
Populations of all sorts of organisms will show marked differences in the course of time. Seasonal variations are regular, but unexplained differences are also observed. In fact, population studies were among the earliest investigations to show the phenomenon that became known a few decades ago under the name of “chaos theory” or non-linear dynamics. Studies by investigators like May and Verhulst revealed the “bifurcation graphs” that showed how the size of successive populations may show huge differences without any obvious cause being discernable. A sudden major difference in the abundance of a diatom species need not indicate a major ecological change, therefore, but if the populations of all diatom species disappear — as in the present case — something is certainly wrong! As diatoms are one of the principal foodstuffs for many animals, the effect of such an eclipse will propagate upward through the food chain.
That recovery took place during the winter months nicely demonstrates how many diatom species do not mind the cold. On the contrary: numerous diatom species rank among the principal cold-loving “extremophilic” organisms, constituting the bulk of the massive plankton blooms in polar seas, for instance. Also, there is a wide range of typical “ice-diatom species” that grow on — and even within — sea ice in the polar regions, and finally, rich crops of freshwater diatoms are seen growing on the lower ice surface when ditches or lakes have been frozen for some time. The cold-loving extremophilic species produce special “ice-binding proteins” and a substance bearing the lovely name of dimethylsulphoniopropionate (DMSP) that protect the organism against damage from the freeze-thaw cycles. Hardy critters, the diatoms!
All comments to the authors Michael Stringer and Frithjof Sterrenburg are welcomed.
*****
Microscopy UK Front
Page
Micscape
Magazine
Article
Library
Published in the January 2011 edition of Micscape Magazine.
Please report any Web problems or offer general comments to the Micscape Editor .
Micscape is the on-line monthly magazine of the Microscopy UK website at Microscopy-UK .
© Onview.net Ltd, Microscopy-UK, and all contributors 1995 onwards. All rights reserved. Main site is at www.microscopy-uk.org.uk .