
|
Puzzles, Enigmas, and Perplexities: (Part 1 of an Infinite Number of Parts) by Richard L. Howey, Wyoming, USA |
The other morning, I woke up still puzzling about sponge spicules and it occurred to me that if I had one year to investigate each thing that I am perplexed about, then I would live forever. I’m afraid I was born with a superabundance of curiosity which has created, over time, a number of awkward moments but, even so, I can’t understand people who have so little curiosity or interest in the extraordinarily enigmatic and magnificent universe in which we live. I am possessed by that terrible god-like passion that obsessed Faust: “Zwar weiss ich viel, doch möcht ich alles wissen!” (“Indeed, I know much, but I would know everything!” However, no sudden universal epiphany when I become omniscient; I want this to occur day-by-day–a constant struggle to discover and learn the incalculable number of secrets of nature.
As a consequence of these early morning musings, I decided to construct a brief “catalog” of puzzles about invertebrates and microscopy and, out of concern for my fellow non-immortal humans, I shall make a Herculean effort to keep it down to 2 or 3 essays.
Since I have recently been rambling at some length about sponges, thatseems a good place to start.
1) To me, the variation in the types of spicules is dazzling and, on this subject, my main puzzlement is about the cells which produce them.
a) Are there a lot of different sorts of cells that have genetic programming that determines that each sort will produce a particular type of spicule?
b) Or is there a highly versatile type of cell (like a version of a “stem cell”) which can respond to specific environmental sponge location-specific “triggers” to produce a particular type of spicule or basal strand or fused process for the osculum?
c) What determines whether there is a fusing of spicules in some cases or organic links connecting spicules?
d) How important are spicules in the taxonomy (classification) of Porifera (sponges)? In 1978, the late Dr. Patricia R. Berquist, a research specialist on Porifera, in her books simply titled: Sponges, wrote : “There is nothing more boring than descriptions of the diversity of spicule forms in sponges, and in the final analysis nothing so misleading as a classification which leans too heavily on such descriptions.” (p. 91) Well, after such an heretical remark, had Professor Berquist ever applied for membership in our group SLOP (Spicule Lovers of Porifera), her application would have been refused. So, permit me to “bore” you briefly with a few of my own images and some URLS which show that sponge spicules are anything but boring and in taxonomic research, descriptions and images should complement each other. Sponge spicules images.

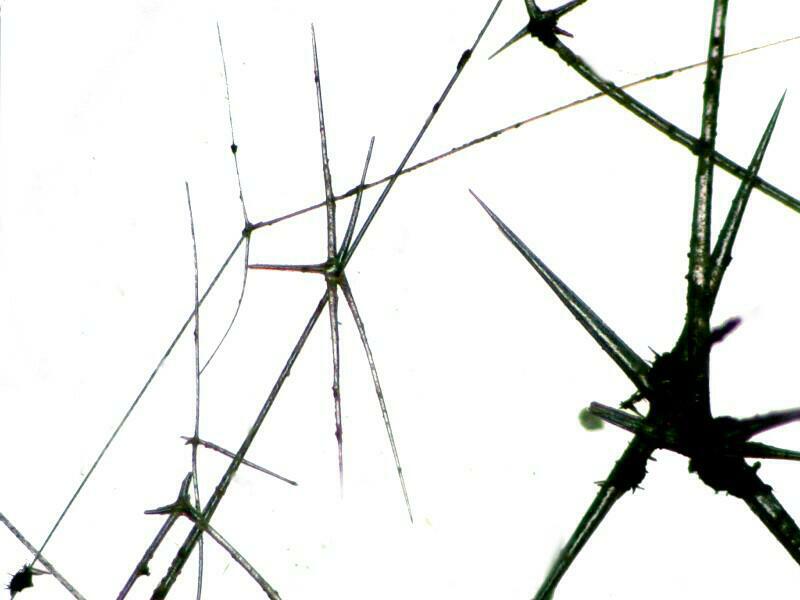
2) Let’s move on to some other types of structures, the “shells” of radiolaria which are also composed of silica although a closely related group, the acantharia have “shells” composed of strontium sulfate. Radiolaria are extremely ancient organisms and there are enormous deposits of fossil shells. They are very small–a tiny sample in the bottom of a small vial can contain thousands of specimens; they are exclusively marine and they have an architectural elegance that is unrivaled. A major deposit of fossil shells is found off the coast of Barbados and it is interesting to note that these Caribbean forms can vary significantly from forms found in the Pacific. Radiolarian ooze is often a combination of radiolaria, silicoflagellates, acantharians and diatoms. As these organisms die in the upper water layers, they gradually descend taking weeks or even months to reach the bottom and at certain levels there is an etching and even dissolving of the opaline silica. This means that in some cases, we may have a very misleading notion of a particular species if we are limited to examining only a very few specimens of that type. When people first learn about radiolaria, they are usually surprised to discover that these tiny aesthetic marvels are formed by amoeboid organisms. What hubris! How can lowly amoebae aspire to creating such elegant architecture? Well, they manage and with remarkable variety and complexity as you shall see. By microscopic standards these shells are relatively thick and combined with the fact they are largely composed of silica (glass) that makes them a challenge to photograph using traditional optical microscopy (but it can be done). They are, however, wonderful candidates for SEM photomicrography. Radiolaria images.


3) Almost everyone who has ever examined pond water with a microscope has encountered that marvelous little organism called Vorticella which was first described by Leeuwenhoek. It is shaped like an inverted bell with a ring of cilia around the outer margin which has sufficient intensity that one can observe tiny flagellates and ciliates caught in the vortex of the current and swept down into the cytostome. Furthermore, it is on a stalk 5 to 15 times the length of the bell, but even more amazing is that fact that the stalk is contractile and if something bumps up against the Vorticella or if it detects a toxin, it quickly contracts by both coiling the stalk and closing the top portion of the bell.
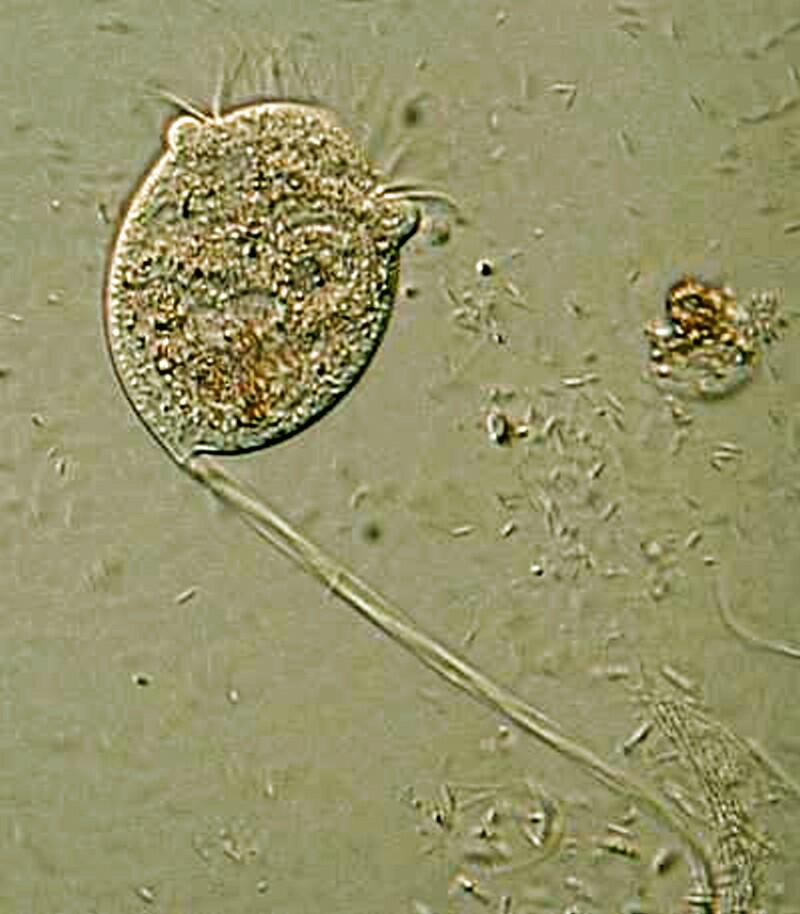
If you look carefully at the stalk, you can see a thin fiber running through it which is the myoneme (or spasmoneme) and it functions like a micro-muscle controlling the contraction. Then, as the threat passes, it seems like the myoneme relaxes and the stalk uncoils, the bell opens up again and the double ring of cilia become active once again. However, if a stimulus combines to make the Vorticella retract repeatedly, the reaction gradually becomes slower suggesting a complex biophysical-biochemical process rather like the gradual weakening of the discharge of a battery.
A puzzle (and I admit that I am easily perplexed) is that this organism can construct this quite sophisticated mechanism of a contractile stalk and this suggests some impressive genetic engineering here.
However, as the saying goes: “You ain’t seen nothin’ yet!” Vorticella has 2 fairly close relatives, both of which are colonial. They have inverted bells on stalks that are attached to other stalks on a central stalk. The 2 organisms are Carchesium and Zoothamnium and the difference between the 2 is quite extraordinary. In Carchesium if one or more of the zooids (stalked bells) is disturbed it (or they) will contract. However, in Zoothamnium if there is a threat or disruption, the entire colony contracts! Here we see an example of Mother Nature (or Big Mama, as some of my students used to call her) experimenting wildly as though testing out the relative advantages of individual zooid contraction as against the “automatic” contraction of the entire colony. Well, both strategies seemed to have worked well because, after millions of years, the descendants of both of these organisms are still around.
Here is a site which shows you examples of Zoothamnium.
And here is another site showing you images of Carchesium.
The basic puzzles here are fairly obvious: a) How can Carchesium be so selective (in some cases) that only one or a few zooids contract? and b) Why does Zoothamnium contract collectively as though it were a single organism?
4) While we’re discussing stalked protists, we need to consider Anthophysis. Imagine a micro-tree which about 1/3 of the way up the trunk begins to branch in a moderate fashion. Further imagine that at the tip of each of the branches, there is a round, fuzzy-looking sphere which is one of the most extraordinary features of this bizarre organism. However, let me systematically list my mini-catalog of puzzlements with regard to Anthophysis. Here is a URL which provides a video clip. The first 1 minute and 20 seconds is definitely Anthophysis, however, the subsequent material which shows deep green colonies is, I suspect, a different organism.
http://www.youtube.com/watch?v=-z3odIzGMs0
In almost all of the specimens I have examined, the stalk is a brownish color. In part, this is not such a puzzle, since these organisms tend to flourish in iron-rich waters. As iron is absorbed the stalk darkens and we can observe the same process in the “shell” of the amoeba Arcella in which the depth of color is an indicator of the age of the organism.
However, what I am curious about is the structure of the stalk of Anthophysis, how it grows, and how it branches. The answer would depend upon doing a series of thin sections using a very good microtome and, in addition, having rich, stable cultures so that you could spend hundreds of hours observing the whole living colonies while, at the same time, trying to figure out what happens to the sub-colonies when they separate and go spinning off. For the amateur, the equipment and techniques necessary to deeply investigate such issues are beyond reach, nonetheless, I remain curious. The most tempting analogy in terms of approaching an understanding of this organism is that of a small tree. There is a central stalk or trunk, branches, and at the end of the branches, leaves or, in this case, compact cluster more like a flower. At the tip is spherical grouping of tiny flagellates each of which have 2 flagella and when the larger colony is disturbed–as it is when you pipette it out of a culture and onto a slide–this sphere may separate off from the tip and go rotating off. So, here’s another puzzle: does it in its natural environment go off and attach, secrete an anchor which becomes a stalk and begin producing a new supercolony or does it simply disintegrate? Another possibility is that some of the individual flagellates separate from the cluster, anchor themselves, and start a new colony. There are, after all, a number of stalked flagellates (some single, some in groups) and also some stalked ciliates (again some single, others colonial) such as Vorticella, Epistylis, Zoothamnium, not to mention the very weird suctorians. To me, it would seem that these processes of producing branching stalks would be rather different in the flagellates, ciliates, and suctoria than in groups such as hydroids, bryozoa, and colonial tunicates where, in many instances. the colonies are extended by a process of budding. The problem is that very little is known about these phenomena and very little comparative work has been done.
3) Volvox, especially V. globator or V. aureus, is an organism which every amateur microscopist should examine live specimens of; these are fascinating, elegant, and surprisingly complex organisms. To get a sense for these remarkable creatures, go to Wim van Egmond’s excellent article. Reproduction can be sexual or parthenogenic. The “individuals” which are embedded in a gelatinous matrix possess a stigma (eye spot) and pyrenoids for storing starch.
Way back at this “primitive” level in the development of the biological kingdoms, naughty Mother Nature was already experimenting with variant sexual types. Volvox is, microscopically speaking a large transparent spherical colony consisting of a few hundred to a few thousand cells, each of which have 2 flagella. Within the large sphere, small spheres (subcolonies) develop and in some, all of the subcolonies will be female, in others, all male; and in yet others a mixture of male and female subcolonies.
The structure of this organism is astonishing. On the outer surface of the sphere, we find the “individual” flagellates embedded and, by the way, each contains chlorophyll so they manufacture their food photosynthetically. Each “individual” has its own little hexagonal chamber in which it resides but, there are delicate fibrils which provide interconnections with neighboring chambers. This suggest a sort of network reminiscent of the Borg, for you Star Trek fans. So, now we have these thousands of flagella stirring up the water and the colony gently spins through the water in a marvelously elegant fashion. And here’s the puzzle: How is the action of these thousands of flagella coordinated? Imagine the chaos if half of them were beating in one direction and the other half in the opposite direction. There certainly isn’t any central processing unit, no nervous system, nothing to coordinate all of this activity and yet, coordinated it is, just like a Busby Berkeley water ballet. However, the “cells” in Volvox need no training; there are, so far as we know, no sensors signaling to each other and yet everything comes together in a smooth and efficient fashion.
Up through much of the 20th Century, the temptation was to offer a mechanical explanation for such phenomena. I can imagine some early 20th Century Rube Goldberg engineer constructing a mechanical, beautiful, polished copper model that had been treated to acquire a striking green patina. It would possess thousands of tiny clockwork gears which would make the 2 filaments (mechanical flagella) in the thousands of units vibrate in harmony when the mechanism was wound up. Why not? Look at the extraordinary singing mechanical birds in cages which the Swiss produced. However, even if one could produce such a model, it still wouldn’t solve the problem of coordinating the filaments in a fashion that would produce spherical rotation.
By the middle of the 20th Century, the biological sciences, aided by advances in computer technologies, were beginning to make quantum leaps forward. The new insights provided by molecular biology, bioengineering, nanotechnology, materials sciences, and the refinement of analytical observation technologies, such as, transmitted and scanning electron microscopy, field ion microscopy, atomic force microscopy and laser confocal microscopy all contributed to providing a basis for new forms of conceptual modeling that go beyond the traditional, intuitive mechanical formulations.
We are now in a somewhat better position to think in terms of dynamic models that can modify and “improve” themselves. The reason for the qualification regarding improvement is my anti-teleological disposition, that is, change does not necessarily mean improvement which is why so many people misunderstand both evolution and politics. The challenge of learning how to develop and think in terms of new conceptual models is a task of Brobdignagian proportions. Think of how many centuries it took for the heliocentric view of the solar system to supplant the geocentric perspective and there were real intellectual casualties along the way–Copernicus, Giordano Bruno, Galileo and others who are now barely remembered. Newton tried brilliantly to console us with an harmonious view of the cosmos intricately regulated by Absolute Laws. But then there were a bunch of spoilsports like Einstein, Bohr, Planck, Dirac, etc., who not only threw relativity at us, but indeterminacy as well. It’s enough to make one check into the nearest monastery or convent, unless you happened to be one of those cursed beings whose curiosity cannot be contained, in which case you check into your nearest university or research laboratory and are discouraged to learn that many of the investigators believe that the foundation basic principles are in place and now it's just a matter of filling in the details. This is the disposition which I call hyper-hubris. If, in a few centuries we’re still around as a species and still doing science and developing new technologies, we will look back at our present arrogance and be appalled. There are already examples that should give us pause regarding our “quasi-omniscience”, the fact that we cannot efficiently sequester carbon dioxide to keep it from destroying most higher life forms on the planet and the fact that we have not yet even identified more than about half of the organisms with which we share on Earth.
A much more modest example is the fact that I went on the Internet this afternoon to search for information on the locomotion of Volvox. (See, I hadn’t forgotten what we were talking about.) I was enlightened with the fact that it is a colony which is propelled by flagella. Admittedly, my search was cursory and somewhat hurried but, there wasn’t a single reference to the issue of coordination. I’ll do a search in greater depth later on. Of course, this is the kind of research that is very difficult to get funding for as against projects to develop more sophisticated surveillance and killer drones.
It occurred to me that it might be easier to investigate the issue of coordination in simpler colonial flagellates, such as, Gonium, Pandorina, Eudorina, or Synura where the number of individuals range form 4 to 64 individuals. It would seem that with a colony of just 4 units, things should be simpler and more evident. (If we say “simpler”, why can’t we say “evidenter”? Stupid irrational language! It must have been devised by a bunch of drunks.) Again the temptation is to rely on a mechanical model; we have 4 units, each has 2 flagella, they beat back and forth and the colony moves. What’s the problem? Perhaps, at this scale, even if the flagellar movement is random, it is limited enough that it doesn’t matter. Whatever happens, they produce motion. Nature kept experimenting and there are colonial flagellates consisting of 8, 16, 32, 64, 128 individuals culminating in the large Volvox which as we noted, can have as many as 50,000 individuals. Perhaps here, a simply mechanical explanation does suffice with smaller colonies but, once we get beyond a certain order of magnitude, it is difficult for me to envision how all of these individuals’ flagella work together to achieve the desired result. It’s like trying to envision 50,000 soccer fans coordinating their behavior after losing a world match. However, Ialso think that it’s a complete waste of time to look for some central control structure.
Let me give you a parallel case with regard to ciliates, such as, Paramecium. It has long been known that groups of cilia in such organisms move in patterns called synchronous waves. In the 19th Century, a number of prominent protozoologists proposed the theory of a central control structure called a “motorium”. This was thought to be the “mechanism” which accounted for this marvelously, intricate series of patterns that guided the motion of paramecia. There are elaborate explanations, drawings, and even some photographs of specimens, using special staining techniques, all designed to demonstrate the existence of the “motorium”. If one does a search of current literature on this subject, one finds only a few passing references and nothing that suggests that anyone is still committed to this theory.
This leads me to a brief but fundamental important digression. I strongly suspect that what earlier microscopists described as a “motorium” was an artifact, that is a pseudo-structure, which was the result of killing, fixation and/or staining techniques. Artifacts can be extremely difficult to root out–use the same technique over and over and you get the same pseudo-structure over and over. Years ago, I was awarded an Arts and Sciences Professorship to work for a year in a different department. (I always thought that this was a plot on the part of my colleagues in Philosophy to get rid of me.) I chose to work in the lab of a distinguished colleague in Zoology, who was a first rate protozoologist, cell biologist, electron microscopists, and director of the Center for Microscopy, Robert A. Jenkins. We worked on that wonderfully strange ciliate, Lacrymaria olor which can extend its neck up to 10 times is body length and, at the time, he was also working on the ciliate Spirostomum minus, a highly contractile organism which I had originally supplied to him and periodically I had to go out and find again for him, since his lab assistants were not too scrupulous about maintaining the cultures. Fortunately, it is an organism which cultures readily. He was doing ultra thin sectioning especially of the chain nucleus and examining them with a Transmission Electron Microscope. One of the first conversations which we had that year was about the problems involved in interpreting an image at every level from ordinary light microscopy through all of the contrast techniques and on up to the highly sophisticated techniques involved in SEM images. He and I shared a delight in observing and investigating these organisms which led to a strong desire, whether dealing with behavior or morphology, to “get it right”.
So, if there are no control coordinating structures in Paramecium, Spirostomum, Blepharisma, or Volvox, then how do we account for their remarkably complex behavior. Give me an SEM a TEM, an ultramicrotome, a sputter coater, 5 lab technicians, a laser confocal microscope, digital cameras for all of the above, a building to house the above, adequate maintenance, salary, supply operating funds and 2 vending machines–one for single malt Scotches and the other for first class gins–and 19 years and I’ll give you an answer. It would probably be the same answer that I’d give you right now if you pressed me for an explanation–“I don’t have a clue.” But, think of all I would have done to stimulate the economy.
Seriously, one of the barriers in our finding answers to such questions is our reflexive approach, we suffer from a kind of cultural conceptual disability. It is a virtual cognitive reflex that when we notice complex behavior, we begin looking for a coordinating mechanism and/or network; something like a brain and a nervous system and then we can say to ourselves: “Oh, that’s how it works” even though we may have almost no clue regarding how a particular brain or neural system may function. In many instances, if there is something we can point to which we believe to be the coordinating “agent” along with a set of connections to the various functioning parts, then that is psychologically often enough for us to rather blithely assume that we have, at least in principle, an explanation.
What is befuddling to us is that things that are so “simple” can be so enormously complex and capable of behavior so sophisticated that we find it almost incomprehensible. Among other recent disciplines, nanotechnology is teaching us some of these very basic and startling lessons. With the sorts of instrumentation that we have developed over the last 75 years, we are in a position to begin poking and prodding Nature in ways never before imaginable. However, we need to remember how rudimentary our understandings and explanations still are and not let our egos get inflated with the bits and pieces we have unraveled thus far. e.e. cummings expresses these cautions quite eloquently in the 2 poems which you will find below.
O sweet spontaneous
earth how often have
the
doting
fingers of
prurient philosophers pinched
and
poked
thee
,has the naughty thumb
of science prodded
thy
beauty ,how
often have religions taken
thee upon their scraggy knees
squeezing and
buffeting thee that thou
mightest conceive
gods
(but
true
to the incomparable
couch of death thy
rhythmic
lover
thou answerest
them only with
spring)
------------------------------------------------------------------
pity
this busy monster, manunkind,
not.
Progress is a comfortable disease:
your
victim (death and life safely beyond)
plays
with the bigness of his littleness
--- electrons deify one razorblade
into
a mountainrange; lenses extend
unwish
through curving wherewhen till unwish
returns
on its unself.
A
world of made
is
not a world of born --- pity poor flesh
and
trees, poor stars and stones, but never this
fine
specimen of hypermagical
ultraomnipotence.
We doctors know
a
hopeless case if --- listen: there's a hell
of
a good universe next door; let's go
Who would have thought that the cilia of a Paramecium would have the same basic morphology as the cilia in human lungs? We can now also examine and describe the structure of tiny fibrils which are capable of a gliding and telescoping behavior that helps us begin to understand phenomena such as contractility. Nonetheless, we are still surrounded by endless, elegant puzzles.
In Part 2, we’ll return to some further mysteries including those regarding Volvox.
All comments to the author Richard Howey are welcomed.
Editor's note: Visit Richard Howey's new website at http://rhowey.googlepages.com/home where he plans to share aspects of his wide interests.
Microscopy UK Front
Page
Micscape
Magazine
Article
Library
Published in the March 2013 edition of Micscape Magazine.
Please report any Web problems or offer general comments to the Micscape Editor .
Micscape is the on-line monthly magazine of the Microscopy UK website at Microscopy-UK .
©
Onview.net Ltd, Microscopy-UK, and all contributors 1995
onwards. All rights reserved.
Main site is at
www.microscopy-uk.org.uk .