|
|
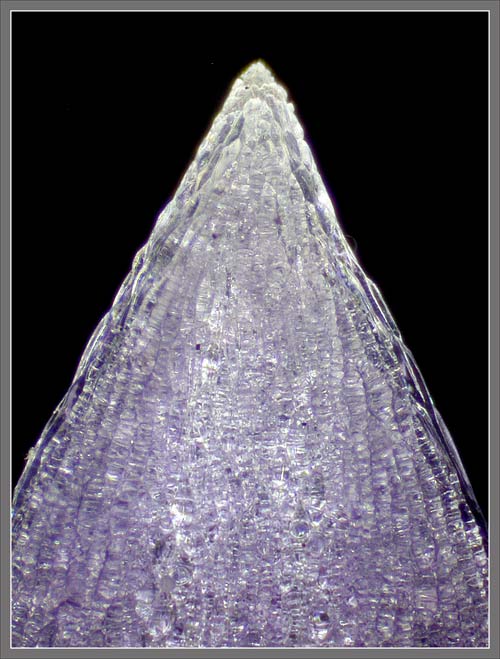
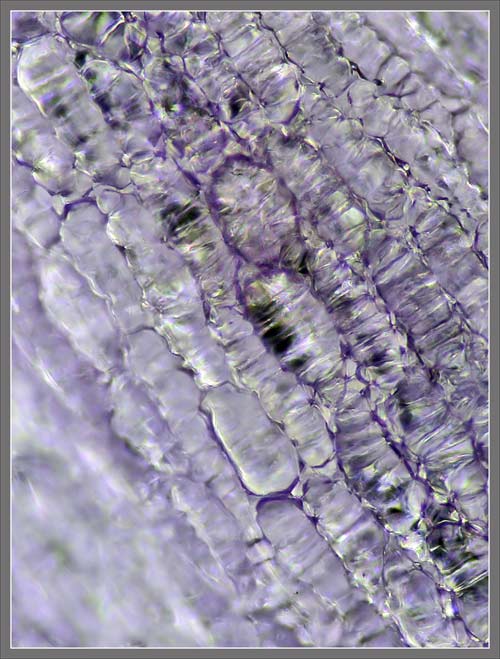
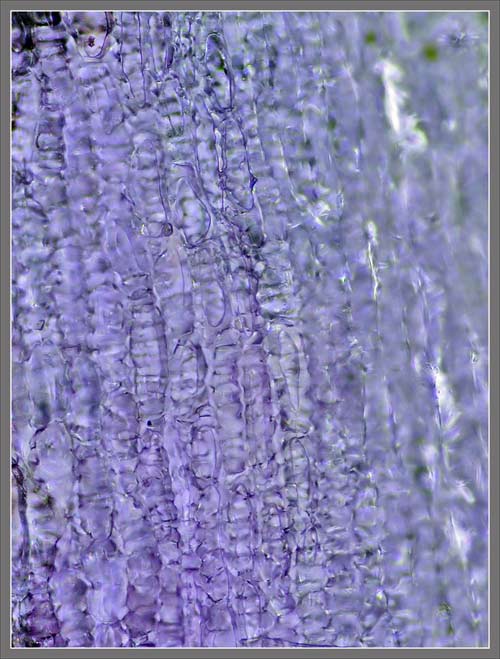
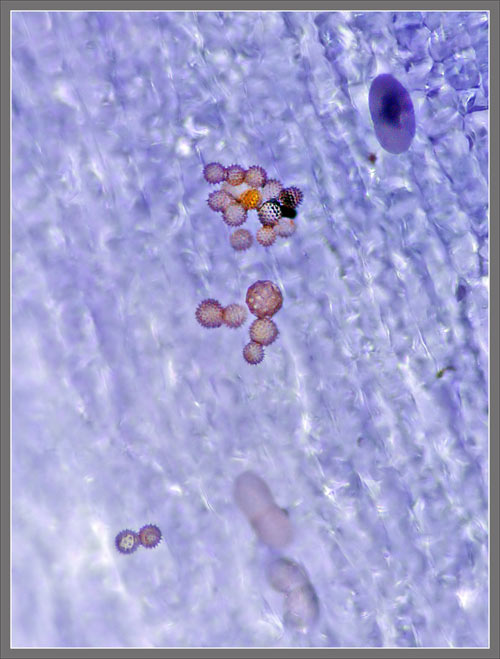
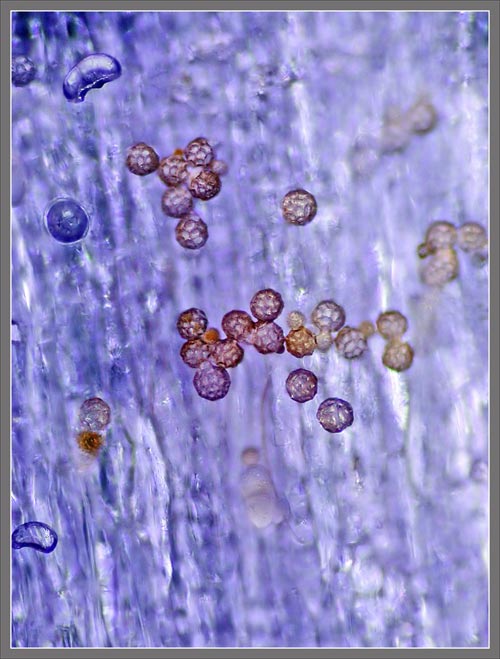
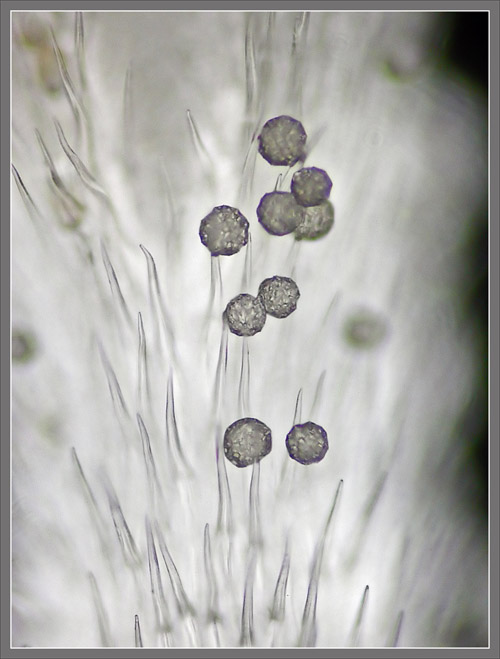
A
Close-up View of a
Stokes' Aster Hybrid
|
|
|
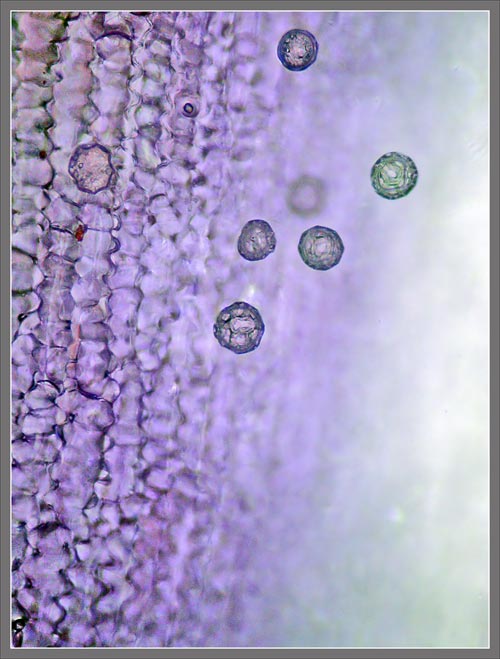
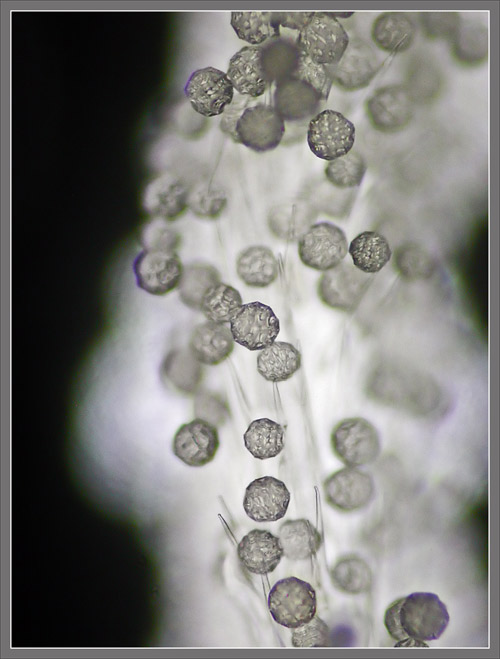
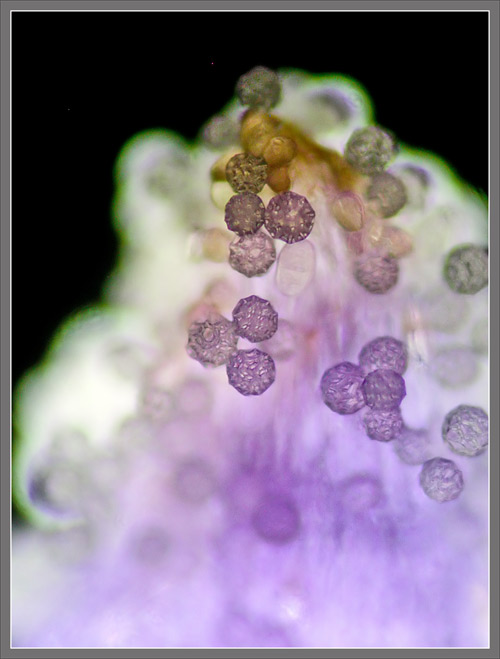
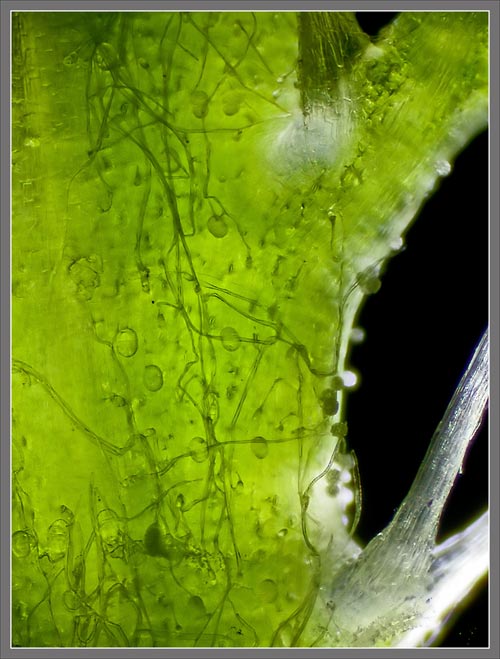
A
Close-up View of a
Stokes' Aster Hybrid
|
































































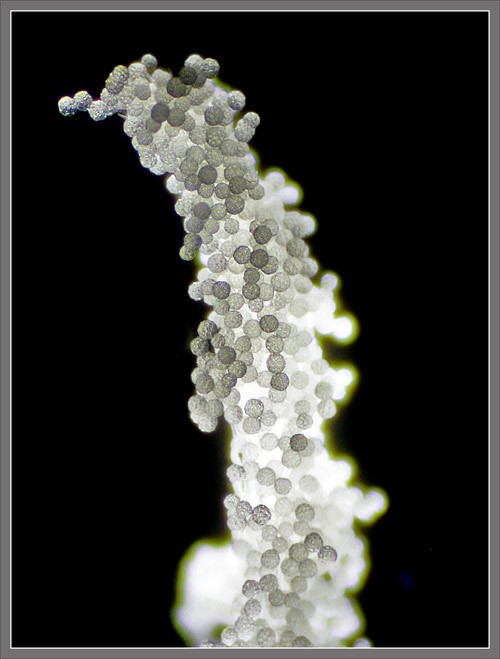
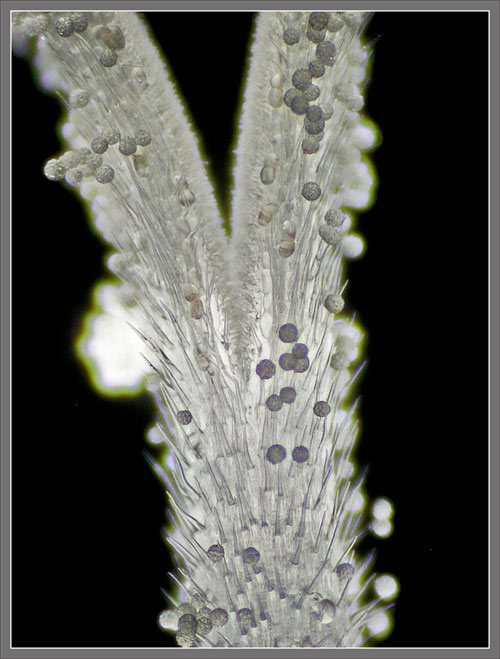

























Published in the
November 2009 edition of Micscape.
Please report any Web problems or
offer general comments to the Micscape
Editor.
Micscape is the on-line monthly magazine
of the Microscopy UK web
site at Microscopy-UK
© Onview.net Ltd, Microscopy-UK, and all contributors 1995 onwards. All rights reserved. Main site is at www.microscopy-uk.org.uk with full mirror at www.microscopy-uk.net .
