 |
THE REPRODUCTION OF VORTICELLA. |
|
|
|
THE REPRODUCTION OF VORTICELLA. |
|
|
The "vorticella" are attractive, quite abundant organisms, and as with many species, most of them cosmopolitan. They are common and easy to observe (a Vorticella was the first protozoan described by Anthony van Leeuwenhoek), but difficult to prepare if you want to study their cytology.
They are bacteriophages, which allows amateurs to cultivate them in the laboratory, introducing into the container in which they were found, small amounts of organic matter (e.g. portions of cooked cereal grains, cereal food for babies) to obtain the development of a bacterial population, but taking care that fermentation stays at low levels. Few species live in really putrid media.
They can be found on almost any submerged substrate, but especially on the leaves and roots of the aquatic plants, or on crustaceans (especially crabs) with large surfaces that cannot be reached for cleaning by the legs and claws, or on shells of gastropod molluscs.
Until the development of electronic cameras, that allow us to obtain a series of fast successive pictures, the reproduction of vorticella had to be taught by means of drawings.
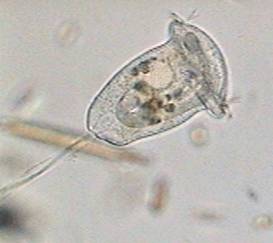
Anatomy of a Vorticella

Here we have a whole specimen, enveloped in its transparent superficial pellicle (ps) finely striated crosswise. Although not excessively sharp, the picture allows all the important organelles to be distinguished, except for a small spherical body: the micronucleus (that really can be any one of the several visible in the cytoplasm) which is a compact center of all the genetic information of the organism. The fact that the macronucleus (mn) with its typical form of a band curved like a horseshoe is so visible in this species is a gift that will allow us to follow with more detail the reproductive process.
Of course everyone that has seen a vorticella know that its peduncle (p) is contractile. The contraction is caused by the "spasmoneme" (sp), the wavy filament that is seen in the interior of the peduncle. It was denominated "myoneme" (muscular filament), before it had been discovered that the chemical system that administers the contraction of the spasmoneme is different from which a true muscular fiber uses. The attached animation (that was captured and adapted for the screen by Jean Marie Cavanihac, who has kindly allowed its use in this article) shows the typical contraction of the peduncle of a species of vorticella.
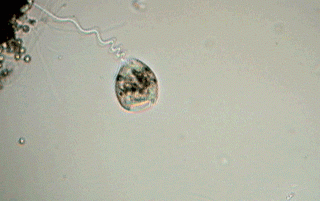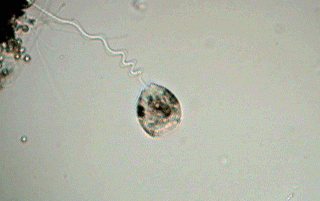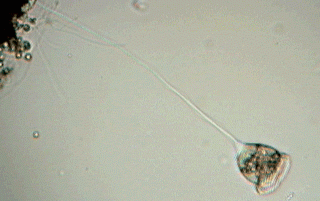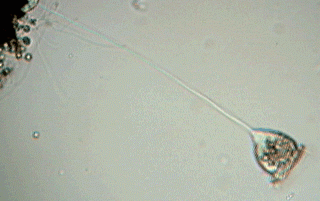
Some microfibers (mf) of the spasmoneme (that can be seen in the picture) enter the zooid (the body of the vorticella) and are perhaps part of the system that retracts the apical disc and the ciliated ring.
The ring, which is more of a spiral than a ring, has 3 rows (membranelles) of cilia (m1, m2, m3) of great mobility, which cause an eddy or vortex in the water (from which the name of the genus was derived) that attracts the food towards the, in this case, small buccal funnel (bf). At the end of the same structure, digestive vacuoles (dv) form successively for the animal to capture and digest their food (bacteria and very small flagellates).
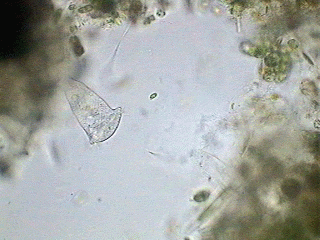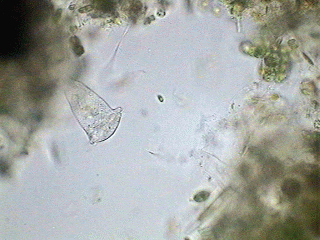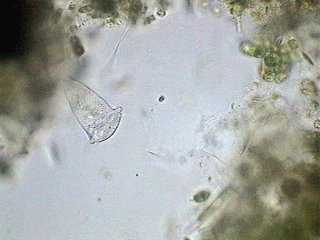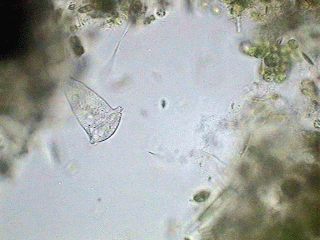
In the anatomy picture shown earlier can be seen the circuit of the vacuoles, first downwards, where they are accumulated and next towards the left and above. The short attached film above shows the typical feeding activity of Vorticella.
At some point in the cell, near the upper rim, is located the cytopige, a fixed site, to which the vacuoles with the digestive remains approach, and where a temporary orifice is opened suddenly to expel the non digested remainders. It requires time and opportunity (with good presentation and little mobility of the zooid) to be able to observe this process.
The liquid remainds, which contain salts that must be expelled to maintain the internal balance of the cytoplasm, are accumulated in a contractile vesicle (cv). A short time of observation of a live vorticella allows this system to be seen in operation. The full vesicle abruptly contracts and seems to disappear, only to return to fill more or less slowly in the same place.
The size and aspect of the species shown, especially the form and position of the macronucleus and the size and disposition of the digestive vacuoles, suggests that it is Vorticella convallaria. A fresh water species alike V. similis, which has a smaller size and a nucleus located in a lower position with the vertical part of the macronucleus trailing in the cytoplasm without the upward curvature of convallaria. A very similar species from marine habitats is the very well known V. nebullifera. Many freshwater identifications of V. nebullifera would possibly be attributed to convallaria.
The union between the peduncle and the body is relatively lax and easy to cut. During the observation of a "simple wet mount" it is sometimes possible to see the zooids detach themselves from their peduncle and swim free. This is caused generally by the lack of oxygen in the medium, and the zooids are found accumulated around some bubble of air, or at the edges of the coverslip. Often, before separation of the peduncle, a circle of cilia (the "telotroch") forms at a short distance from the base of the zooid, and helps the zooid in its later free swimming stage. It is common that the name of telotroch is applied by extension to the zooid itself that has the ring of cilia. In his message (number 2176) to the French Yahoo Group MICROSCOPIES, Jean Ma. Cavanihac has presented an image of a detached telotroch (see refs.).
ASEXUAL REPRODUCTION
Only the process of asexual reproduction can be seen in live specimens. All sexual processes in the protozoa implies the activity of the macro and especially of the micronucleus, which display its chromosomes, and interchanges them with another coparticipant. These processes can only be observed by means of fixation and coloration by specific histological stains (i.e. Carmine, Iron Hematoxylin, Feulgen) and mounting a series of individuals in different stages of the process, for its observation obviously at very great powers. This is not really an easy task even for the trained professionals.
Nevertheless asexual reproduction can be observed easily, which makes it all the more strange that on the Web there is no document that illustrates photographically the full partition process of one vorticella. An extensive work, extremely interesting and of easy reading, but illustrated with delicate color drawings has been published by Rosemarie Arbur in Micscape Magazine (see refs.).
As it is seen in this first picture (x10 objective) below, the process begins with a widened zooid, in whose apical disc a vertical incision begins to appear. As this image alerted me I began to periodically photograph the vorticella in division with the x40 objective.
12:27:51
It is typical that the flagellated protozoa reproduce by dividing their body longitudinally, as this beautiful picture below shows. It was sourced from Steven Durr's pages where more pictures on Euglena can be seen.
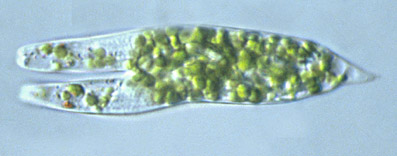
The peritriches (not only the vorticella) are the exception. A registry of the bipartition of an epystilidae one is displayed by Dominique Voissin in his site MICROMEGAS (see refs.).
Click on the following pictures to see bigger ones, taken with the 40x objective.
The
process seen with a x40 objective
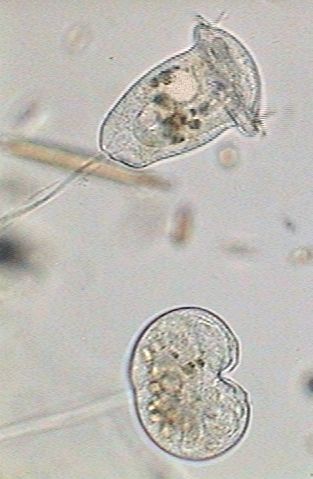
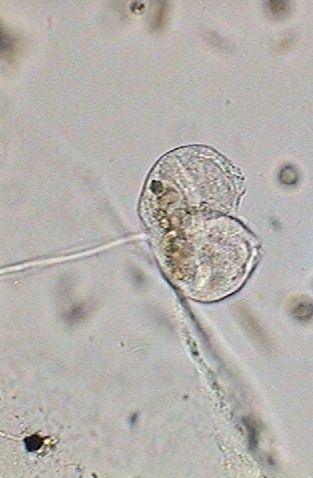
The upper Vorticella in this second picture is the one that has been used for the introduction.
The inferior one shows an incipient vertical split, the accumulation of the digestive vacuoles in the base of the zooid, a long macronucleus extended like a band that crosses the cell, and an involution of the cilia system and the apical disc.
For the rest of the process the vorticella will not be fed and evidently the individual has accumulated food to provide energy for the division.
The nourishing system is closed, with an almost total disappearance of the ciliary activity.
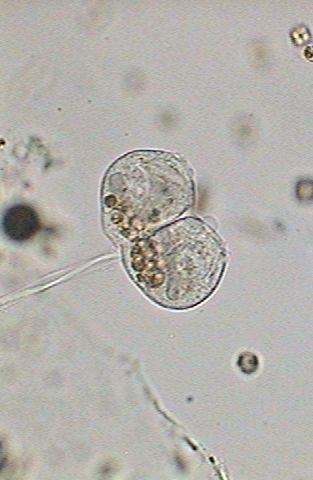
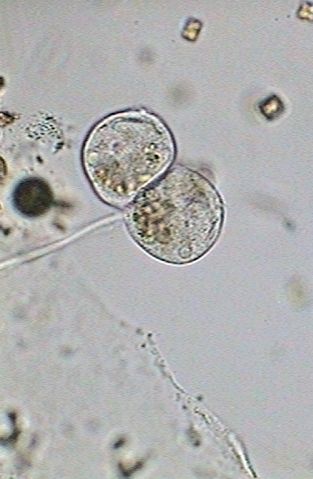
In the
first image (below) the macronucleus has been thinned and stretched. The
numerous nutritional vacuoles are accumulated in the base, in a band that
includes both individuals. Apparently the contractile
vesicle no longer works.
 |
 |
 |
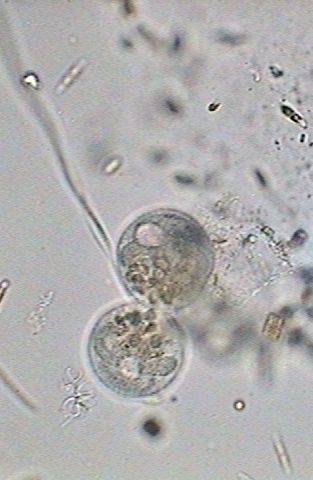
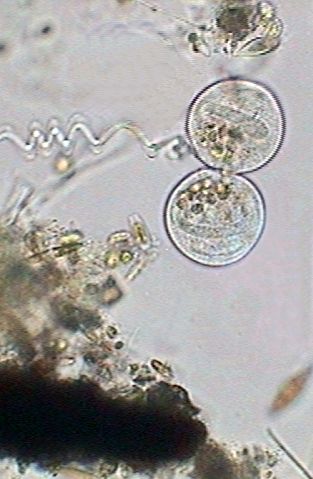
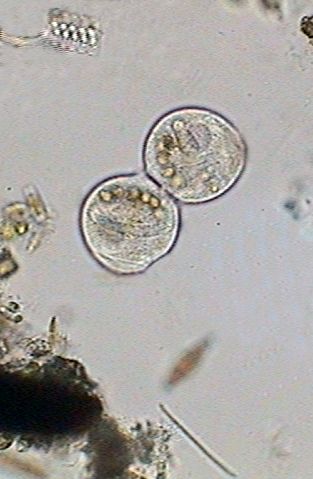
At 12:40:24, in a fast process the macronucleus becomes strangled, divides in two and ceases distributing itself between the two cells (center above). The cilia are mostly immobile and the nourishing vacuoles were distributed between both individuals. The division of the macronucleus implies of course a process of simultaneous division of the micronucleus, that we cannot visually verify by its small size. At this moment (right above) both cells have already an independent macronucleus and a micronucleus.
It recommences
the activity of the contractile vesicle, (see left image below) and still
united between, both zooids leave its peduncle, at 12:59:03 (center image
below). Nevertheless the cilia's activity is still hardly incipient.
 |
 |
 |
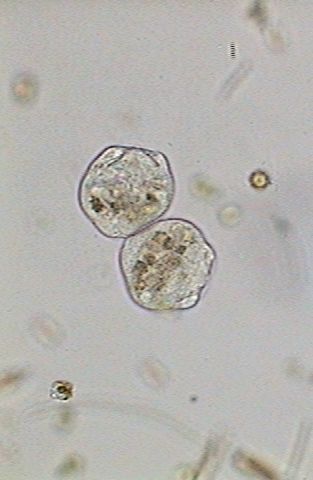
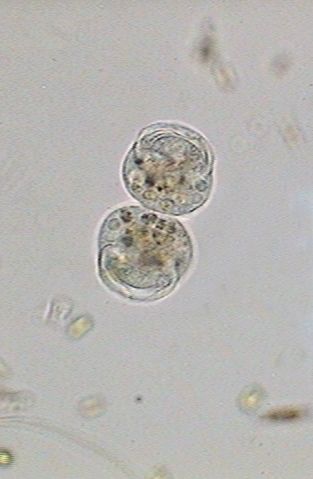
The
ciliary area begins to regenerate (left below), and in the central image
the crown has already opened (13:49:19). The right image shows the beginning
of the differentiation of the "scopula", (13:50:02) the basis of the cell
on whose superior edge the telotroch will be developed.
 |
 |
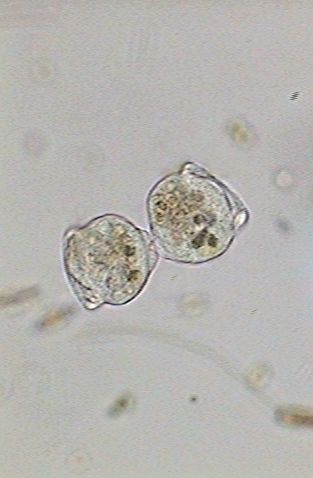 |
From
the moment at which the ciliary activity is reinitiated both zooids are
mechanically working for its separation, because each one tends to advance
in an opposite direction. Nevertheless the maturation of the base of the
cell and the preparation of the telotroch that characterizes the migrant
individuals will take another 19 minutes. See the three images below:
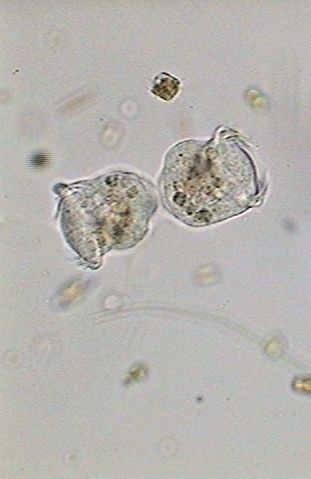 |
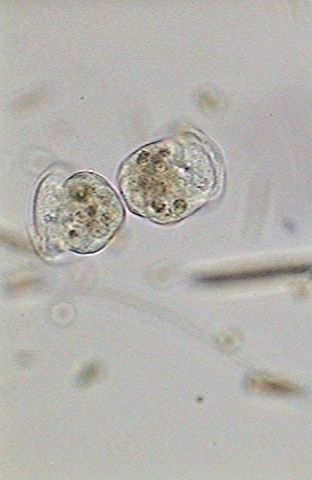 |
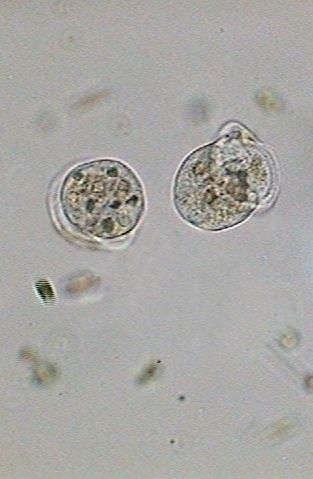 |
| 13:55:11 | 13:56:30 | 13:56:49 |
| The new individuals finish their separation. | ||
The
telotroch, which will develop to maturity like the migratory individuals,
will still take another 13 minutes to complete. See below.
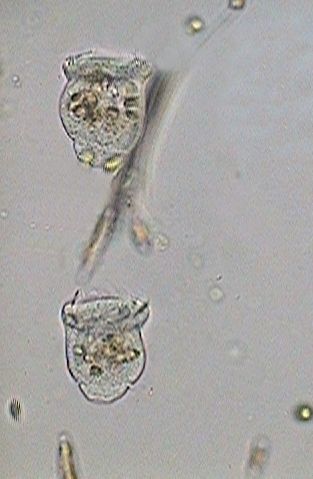 |
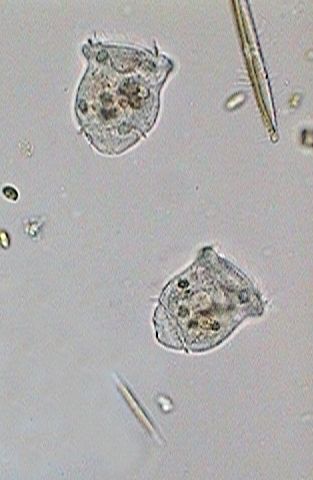 |
Two images above: To 14:05:03 I noticed the first indications of cilia in the telotroch. These were totally developed almost 4 minutes later.
Soon each individual will look for a place that responds to its exigencies of ambience and feeding. On the base of the scopula one can see the zone that will give origin to the peduncle.
Nevertheless, in the surely inhospitable conditions of the confinement between slide and coverslip even one hour later both telotrochs still continued swimming around without making their election. They were put back in the culture media so that they had one better opportunity.
A note that seems important to me is that the reproduction implied a single individual, which was divided in egalitarian form among another two which evidently are not 'father and son', but 'brothers'. The predecessor has disappeared and rejuvenated. This explains the affirmation of many biologists that theoretically the protozoa are immortal.
Comments to the author,
Walter
Dioni, are welcomed.
REFERENCES
ACKNOWLEDGEMENTS
Thank you to Steven Durr for permission to use the image from his web site.
Please report any Web problems or offer general comments to the Micscape Editor.
Micscape is the on-line monthly magazine of the Microscopy UK web site at http://www.microscopy-uk.org.uk with mirror site at http://www.microscopy-uk.net.