
|
Euplectella aspergillum: Part I by Richard L. Howey, Wyoming, USA |
An Essay
Which Examines The Glass Sponge
Also Known
As The Venus Flower Basket
In Respect
Of Its Morphological Aspects
And Also
Looks At Its Social and Symbolic
Significance
in Asiatic Cultures
While
Investigating the General Architecture
Of The
Sponge
With
Supplementary Reflections Upon
The Form Of
Its Spicules Both
In
Individual and Fused States
With Some
Additional Remarks
On The
Creationists' Use
Of This
Particular Organism
To Support A
Highly Dubious Argument
For a Deity
As Designer
Accompanied
By Considerations
On The Issue
Of Individual Organisms
In Relation
To Colonies
I've always wanted to write an essay with a classic Victorian title. [As I worked on this essay, it expanded to such an extent that I shall have to present it in several parts.] I have often been amused by Victorian works whose title and subtitles take up nearly a full page and provide a brief summary of the major themes. I decided that an essay on Euplectella, with its elegance and deceptively fragile appearance, should be a perfect candidate for a Victorian title.
Some time back, I wrote an article which included some observations about the detritus found around the holdfast of Euplectella. I may add a few further remarks regarding the debris, but the focus here will be on the sponge itself and its architecture. I am trying to acquire a couple of specimens from two new sources I have discovered in the hope that they may have been collected from locales different from the specimens which I already have. In such an event, I may find some debris of a different sort which should prove of considerable interest.
A moment ago, I commented on the possibly deceptive fragile appearance of Euplectella. If you drop one on a hard floor, it will likely break; I haven't tried it. However, my suspicions are that it will either a) shatter like glass (which essentially it is) or b) suffer only minor chipping because of the remarkable way in which it is structured—at the moment, with no empirical evidence yet, I am leaning strongly toward the latter possibility. Notice that word "yet." I can hear you asking irately: "What! You're not going to destroy one of those beautiful specimens, are you?" Well, science isn't for the faint-of-heart, so as Bette Davis says in All About Eve "Fasten your seat belts; it's going to be a bumpy ride." (If any of this is inaccurate, blame my wife—she's the movie buff. See Endnote.) So, after I drop one, which will be sometime before I finish the last part of this essay, I'll let you know what happens. Frankenstein that I am, I plan to sacrifice a second specimen to the jaws of a pair of very small bone cutters.
Let me explain this apparent vandalism. The skeleton of Euplectella is remarkably complex and that general fact one can discover without taking one apart. Just looking at the intricate lattice work of the tube is impressive enough but, in addition, there is the series of spiral ridges which coil from the base up around the tube to the top.

Around the rim of the top, there is also a ridge of raised spicules like a balustrade around the roof of some futuristic high-rise edifice. What an extraordinary apartment complex a Euplectella would make—sort of a 22nd Century curving tower of Pisa. Over the osculum (the opening at the top), there is a sieve plate which consists of a complicated lattice of fused spicules. Around the bottom, from an area of 2 inches or so, there arises a series of long, thin glass hairs that partly constitute the holdfast that allows Euplectella to live in muddy, debris-ridden bottoms. It's rather like an aquatic equivalent of anchoring buildings in earthquake-prone areas—the holdfast allows the sponge to move in the currents and diminishes the chances of its being dislodged as the currents shift. Right in that 2-inch area I was talking about, there is, in this specimen, mud in the lattice from this point down, suggesting that this part of the sponge was directly in the substrate. In the previous article called "Cultural Debris" I reported that Twenhofel and Schrock in their 1935 book, Invertebrate Paleontology, mention a conjecture that the mane of hairs anchors Euplectella and that it then more or less floats at a 90-degree angle parallel to the bottom, thereby taking advantage of the food-rich currents. The position of the mud and debris on the specimen I am examining certainly raises questions about the manner of anchoring and I shall keep this issue in mind as I examine other specimens.
I know I still haven't explained or justified my soon to take place acts of vandalism. I'm working up to it, just as it has taken me two years to get to the point where I am willing to sacrifice a couple of specimens. So, this essay is really a work in progress and I will report new facets as I discover them. But why break up such splendid specimens? Because, I have become convinced that there are some things that I want to learn for myself about the wonderful architectonics of this sponge and not simply rely on the accounts, photographs, and drawings of others.
Consider this: sponges, at least from a phylogenetic point of view, are considered to be pretty primitive organisms. They have no brain, no nervous system, no organs; they have relatively few different types of cells; most sponges, for most of their lives, are non-motile and are dependent on the surrounding currents to provide them with food. Nonetheless, these odd collections of cells manage some amazing feats and, as is evident from Euplectella, produce some incredible structures. In fact, it is difficult to know how to talk about sponges. Are they simply collections of cells in a matrix, are they colonies, are they distinct and special organisms? My answer is—yes!—all three! The dominant viewpoint among taxonomists is that sponges descended from a choanocyte ancestor, in other words, from a protist which is a collared flagellate. Such protists still exist today and they are extremely ancient creatures having been around for hundreds of millions, if not over a billion, years. Choanocytes are crucial to the very concept of a sponge, since it is the activity of their flagella that creates the currents through the organism, thus providing food as well as providing a means of dispersing reproductive cells.
Certain specialized cells in sponges are responsible for the formation of spicules or the spongin network of the type found in the ordinary bath sponge. Unless you are a bit of a masochist [the word come from the name of Count Leopold von Sacher-Masoch, author of the infamous novel Venus In Furs], you won't want to massage your delicate skin with calcareous or siliceous sponges and remove a layer two of epidermis. Consider the simplest type of case in the formation of a calcareous spicule which is a simple rod. A minute organic thread is the center of the spicule and a tiny amount of calcareous material is deposited around this axis. This specialized cell is, by the way, binucleate and the formation of the spicules begins in the area between the two nuclei. As the deposition of the calcium carbonate progresses, the two nuclei move further apart and the cell divides. One cell is predominate in defining the length and shape of the spicule and the other, the thickness. In the formation of spicules with three rays, three binucleate cells are involved and each divides, but at the point where the three rays join, those cells remain clustered for a time, depositing layers of calcium carbonate which join the three rays. If there are four rays, then the whole process starts with four binucleate cells. Seems simple and straightforward, yes? Well, no. As it turn out, just in calcareous sponges, there are several dozen different types of spicules and while a few basics are known, many of the details of this process of spicules formation are still unknown.
So, how does it stand with glass sponges and their spicules. It's even more complicated and certainly much more mysterious, since much less is known about the hexactinellids. This term tells us that the spicules have six axes—well, at least a lot of them do. As it turns out, in the hexactinellid sponges, there is also a staggering variety of shapes and types of spicules. So, perhaps, you now understand why my quite intentional and, by no means random, acts of vandalism are necessary. The skeletons of glass sponges turn out to be mindbogglingly complex and they have other types of spicules in the organic matrix which you might not even encounter, since most are lost in the drying and cleaning process, and so far, I haven't been able to get my hands on a preserved specimen that still has the tissue on it.
Let me give you a brief sketch of some of the amazing structures you may encounter if you start looking carefully at hexacintellids. There is, as one would expect, a dense thicket of spicule-type terminology, which you may want to learn if you get really serious about studying sponges. However, I am going to rely on my own naive descriptive approach to attempt to convey an impression of these glass micro-marvels.
1) The simplest sort of spicule is the monaxon or rod. These can be very small or quite long; they can be straight or curved. Seems simple, enough, except that there are all kinds of variations and subtypes:
a) knobbed at one end
b) knobs at both ends
c) a rod, rounded at one end and pointed at the other
d) a rod, rounded at one end and pointed at the other with tiny thorny projections along most of its length
e) pointed at both ends
d) rounded at both ends
g) rods that are strongly curved at the end, so as to form an elongated "C" And, at this point, things get even more interesting, because the hooks at the ends of the "C" can take on a variety of forms:
gl) instead of just a single hook at each end, two recurved hooks at each end
g2) instead of just a single hook at each end, two recurved plates at each end
g3) instead of just a single hook at each end, two recurved flukes at each end
g4) a set of variations where the form at one end of the "C" is different from the form at the other end.
You will recall that I told you that monaxons are the simplest forms of spicules. Sponges are very tricky—going around looking simple and uncomplicated, trying to deceive us. As we look closer and closer into the details of apparently simple organisms and objects, we are almost always in for some surprises. In some respects examining nature is rather like examining music or language carefully. At the most basic level, music seems simply to be a collection of tones. But as we listen and think more deeply, we realize that not only are there while notes and sharps and flats, but quarter tones, and semi-quavers, difference in duration, trills, pizicatti, glissandi, rhythm differences, and, of course, the differences of the "same" note produced by a tuba, a violin, a harp, and an oboe. Then there are factors like tremolo and rubato and suddenly what seemed to be a simple collection of a few sounds becomes an entire universe.
Think what we can do with the 26 letters of the English alphabet; there are estimates that there are over 600,000 words in English. And, of course, there are alphabets in other languages which are radically different from ours and languages, such as, Chinese where there is no alphabet, but rather a complex set of characters based on a set of radicals which can be combined in a multitude of ways.
2) In calcareous sponges, the most common type of spicule is the triradiate, which has three rays or spines and again, this type has a series of variations:
a) rays of equal length at 120 degrees from each other
b) rays of unequal length at 120 degrees
c) T-shaped
d) Y-shaped with straight rays
e) Y-shaped with rays curved upward
f) Y-shaped with rays curved downward
and with c), d), e), and f) the rays may be of equal or unequal length.
3) Tetraxons—As you would expect, these are a series of spicules with 4 rays, usually not in the same plane, which means that they are harder to observe intact under the microscope, for if you put a cover glass on them, there is a good chance of breaking one or more of the rays. This turns out to be important with tetraxons, because deceitful creatures that sponges are, they sometimes form degenerate tetraxons with only 1, 2, or 3 rays and the presence of the "missing" rays can, if you are lucky, be detected only by finding the remains of the axial thread which is organic and may well have shriveled up in dried specimens or deteriorated in poorly preserved ones.
4) Five, do I hear five? Yes, and six, because that's what we've been working up to—six—hexa-hexactinellids. But first, a word about 5-rayed spicules, or pentacts. Imagine a table with 4 supports which rest on the floor, like an "X" and then from the intersection, there arises a fifth rod which is the central support for the table top. Now, imagine another ray extending from that fifth rod and you have a hexactine spicule. So, here we are—we have something that looks like a martial arts weapon, as a sort of basic building block for hexactinellid sponges.
Seems too simple and straightforward, doesn't it? You're right. Glass sponges have almost all of the types of spicules we've mentioned thus far plus some other variants that seem unique to these sponges. Apparently, the only major type of spicule that is absent from the hexactinellids is the spheres and their variants, which are common in another group, the tetractinellid sponges. Oh, yes, there are spheres and some are microscopically speaking, large, others tiny, some are smooth, and others are spiny.
Since the typology of sponge spicules is complicated, let me provide a bit of relief by showing you some interesting of some of the other more common and interesting types of sponges, lattices, and spicules.
First off, is another type of glass sponge which is usually found on Xenophora which is one of the “carrier shells” which you can see here at the bottom.

Next is a portion of the lattice of yet another type of glass sponge and almost looks like a portion of a chandelier.

Sponges are opportunistic in the sense that they exploit virtually any habitat which will provide them shelter and food as exemplified below by the delicate network of sponge growing on a spine of tropical sea urchin.

I mentioned earlier monaxons which is a common and the simplest type of sponge spicule. Here are some monaxons accompanied by several of the “C” type spicules.
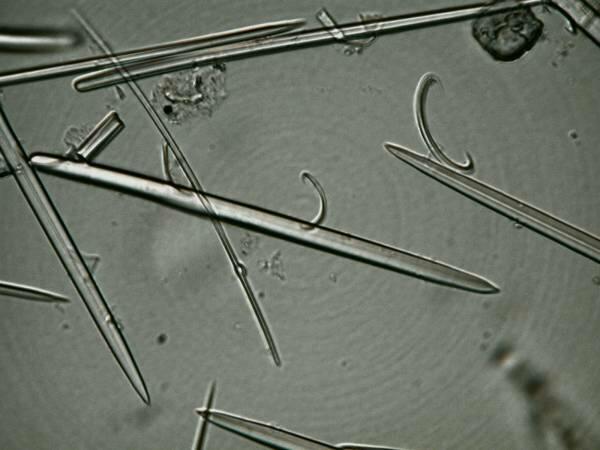
Another type of spicules than can be deceptive simple is the tetraxon and in this case one in the form of a cross.

The image below is of interest for 2 reasons: 1) again it illustrates how sponges will inhabit virtually every possible kind of habitat, since this one was found growing on the surface of a sea anemone and I extracted some of the spicules which are again tetraxons and if you will notice that they are birefringent.
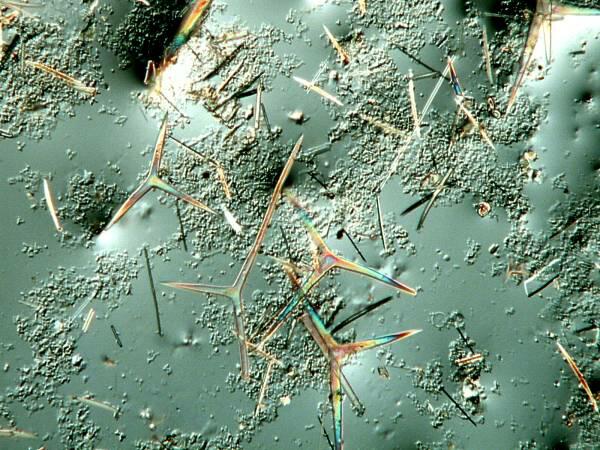
Here is close up of one of those spicules which shows the birefringence even more clearly.

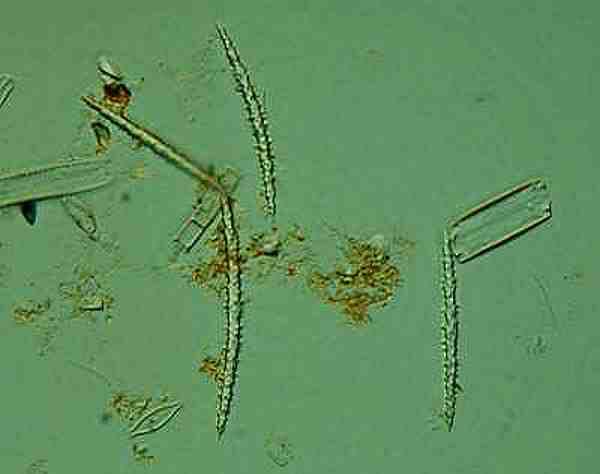
Sometimes monaxons can be thorny and this example is from the freshwater sponge Spongilla (no relation to Godzilla).
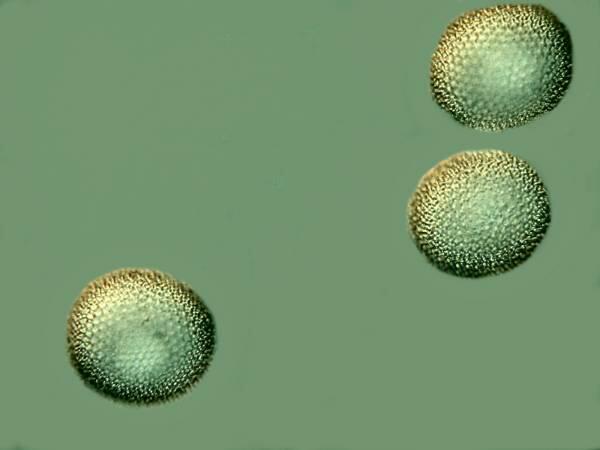
However, as I mentioned before sponge spicules have an enormous variety of types and some of them are strikingly unusual. Here are some examples of some that spheres.

When we look at them at higher magnification, we can see that they are covered with tiny spines.
However, some of the spines belonging to the glass sponges of Hexactinellids, which is the group to which Euplectella belongs, are of stunning complexity. Consider the following example.

Or this spicules, the so-called “dumbbell” spicule, which is from a bizarre sponge called Hyalonema.

Hyalonema is also a glass sponge and I can’t resist giving you a glimpse of it. This one is 5 inches in the longest part of the body, but the glass “tail” is slightly over 20 inches long! It is extraordinary optical qualities which in terms of light transmission exceed our best efforts to produce fiber optics.

Here is a close up of the same sponge and you can see more clearly the mass of glass fibers.

Let me just take a moment to give you an idea of some of the other more intriguing structures you are liable to encounter.
1) parenchymalia—take the basic hexact, but now image 3 points at the end of each tip, so this type of spicule ends up looking like 6 tridents of Neptune joined together. I love the names with spongologists (poriferologists?) have inflicted on us. I fantasize about walking into a market and asking for a pound of parenchymalia or a jewelry store to inquire whether of not they have any pinnulate pentacts ( a sort of feathery spicule which occurs in the genus Hyalonema) or walking into my physician's office to get test results and asking her (yes, my doctor really is female—I'm not just being politically correct!), if the tests showed any pleural prostals, which are spicules that can occur around the osculum or in a very extended fashion as anchoring spicules which have recurved hooks at the tips.
2) If you have ever lived in a 2 (or more) story house and have had to have it painted, then you are familiar with scaffolding. Now, imagine such scaffolding made out of glass spicules which project out in six directions and are fused together to create a wonderful geometric lattice. Further imagine that this lattice itself is the framework around which the house is constructed, that it is dozens of stories high, that it gently curves to form an elegant tube, and that the center is hollow and the roof is perforated. A glass sponge skeleton of this sort is called a dictyonine skeleton.
3) There is a group of spicule types called asters and in their simplest type, they form a sort of starburst with a series of pointed rays extending from a central point. In the hexactinellids, one can find forms that have 6-9 straight bristles at each end instead of a single needle-like point; there are others where each tip looks like a miniature mushroom, and still others in which the tips are so curved and branched that they look feathery.
So, after all of that, you now perhaps understand why I think it's necessary to sacrifice a couple of specimens to investigate further the incredible architecture of glass sponges (and I gave you only the mini-tour).
One can appreciate these sponges just from an aesthetic point of view and marvel at their delicate intricacy or one can be driven, as I was, by that Faustian passion for knowledge. I have learned only a tiny bit of what there is to be learned about Euplectella and only a smattering about the other hexactinellids which produces an immense frustration that helps me understand and appreciate Faust's heretical utterance: "Zwar weiss ich viel, doch moecht' ich alles wissen." (Indeed I know much, but I would know all!)
He wants to be omniscient or, in other words, he wants to be God. Goethe himself was an avid naturalist and his enormous curiosity led him into investigations in optics, botany, and mineralogy. (Goethite is, of course, named after him.) So, I feel rather like Faust—there's so much more I want to know and I can't even obtain specimens of some of the other kinds of glass sponges which I would like to examine.
In the next parts of this essay, I want to examine some broader issues, but ones which will, at least indirectly, link back to glass sponges and Euplectella in particular. However, this part has gotten long enough, but in Part II, I want to reflect a bit on some problems about the relation between colonies and individuals.
Endnote
Although my wife is indeed a movie buff, she is, as well, a patient and long-suffering auditor for my long-winded discourses on everything from termite flagellates to tunicates to pedicellariae. She has proofread every article I have submitted for MICSCAPE and saved me from making some egregious errors. Without her careful reading, my articles would be much less readable and I thank her for all her efforts. (She also told me that if I didn't include this endnote, she was going to stop proofreading!)
All comments to the author Richard Howey are welcomed.
Editor's note: Visit Richard Howey's new website at http://rhowey.googlepages.com/home where he plans to share aspects of his wide interests.
Microscopy UK Front
Page
Micscape
Magazine
Article
Library
Please report any Web problems or offer general comments to the Micscape Editor .
Micscape is the on-line monthly magazine of the Microscopy UK website at Microscopy-UK .