|
The second nematode species I found in
both Durango and Cancún specimens of Periplaneta americana is
HAMMERSCHMIDTIELLA DIESINGI

|
| Schematic like a drawing
of this mature
female shows its pseudocelomate morphology in darkfield. This
is a mosaic of pictures taken with the 10x objective. Click on the
image to see a bigger and labeled version. |
|
The species was
described in 1838 by Hammerschmidt with the name of Oxyuris diesingi. In 1932 Chitwood created the genus Hammerschmidtiella,
making diesingi the type species.
The female, that is described here,
is also a relatively small
nematode, something bigger than Thelastoma, fusiform,
ending in one long, thin and slightly conical tail. It has a short buccal capsule, continued by a triradiate pharynx,
provided with a well differentiated pseudobulb,
connected by a thin but well marked isthmus
to the very muscular bulb (see the
labels in the picture).
The bulb gives entrance to the intestine,
that in these individuals have in their
anterior end one short but defined gastric
expansion (although it does not show any “gastric cecum”) (fig
1 and 2) The rest
of the intestine is straight, thin and long,
finishing in the anus immediately
before the start of the tail (fig
1, 3 and 6).
Fig. 2 - The attached
image shows
a detailed view of the anterior end of the
individual fixed and mounted in 50% glycerin illustrated next (fig 3).
|

|
|

|
Until
2003 H. diesingi was the only species of the genus know to
inhabit
cockroaches. This
year, based on the comparison of genetic sequences of the DNA of the
two
species, a new one was proposed, but not yet denominated according to
my
information, that parasitizes Gromphadorhina portentosa, the
great
sizzling cockroach of Madagascar (which is sold in pet stores, because
there are
many fans that raise and care for it).
Leaving apart the cephalic end,
the cuticle of this species is marked
with very shallow ridges. (Fig.4).
Some
details of the digestive apparatus are in figures 5 - 9.

|

|
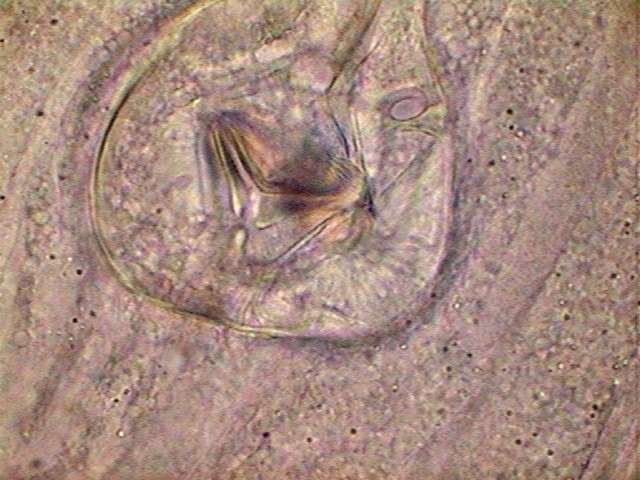
Fig. 5-
Optical section displaying the structure of the mouth and pharynx
|
Fig. 6 -Optical section
that shows cuticle, pseudo bulb and pharynx
|
|

|
Fig
7 - two focus levels to show the structure of the sclerites in the
bulb. They are very different to those from Thelastoma, and may be they
are species specific. Perhaps they could have a role in taxonomy
similar to the trophi in the Rotifera.
|

|

|
Fig 8
- the gastric pouch present in all the individuals.
|
Fig
9 - What I call a duodenum, present in most of the individuals
|

|
Fig.
10 - This is the picture
included in the textbook of Hyman and attributed to the original
description by Chitwood, 1932. There are some differences with our
material, mostly in the structure of the pharynx and the first portion
of the intestine.
|
It
is interesting to note that in the drawing of Chitwood, 1932, the
expansion of the intestine’s anterior end, so evident in our specimens
is not
seen. It seems logical to denominate it
gastric pouch.
Neither the pseudobulb has in his drawing the importance that our
pictures show (see fig. 1,2,3 and 6),
and the isthmus, so evident in my material almost does not appear.
As in
Thelastoma, the strong
muscles that compose
the bulb have sclerites with teeth and grooves in their free edges
which
acts
like tools to crumble the food.
(Fig.
7)

|
Fig 11 . End of intestine.
Colors inverted to better show the muscles and glands of the anus
|
The
Excretory system has an H shape with
short anterior branches, and long posterior ones, like in Thelastoma, that are united
cross-sectionally and end at a ventral
pore
that can be seen in the fig. 1 at
the very end of the gastric pouch.
The
reproductive system is
formed by two thin and long ovaries,
in pictures 1 and 3 in lateral view it is
difficult to identify both organs, but both ovaries are suspected in fig 2 and
they are clearly displayed in the fig. 12. A system with TWO
ovaries is
called
"didelphic". It is seen that, in contrast with Thelastoma, the ovaries both start
at the anterior end (in Thelastoma
one of them was anterior, the other posterior).

|
Fig. 12 - The two ovaries,
both starting at the anterior end, are clearly seen in this picture
|
Nematodes
that have a single
ovary like the species of the genus Blatticola (see the key
at the end) are called "monodelphic".
The ovaries are seen like flexible cords full of small globular cells. Eggs, developed
but not
fertilized, are seen like batteries of coins in the beginning of the
uterus.
Fertilized eggs
enter the uterus, long and with thin
and folded walls,
that finishes in the ventral muscular vagina,
which opens by a small vulva in the
union of
the anterior third with the median third of the body (see figs 1 and 15). In the
older females the uterus can acquire a very large size and be filled
with
hundreds of eggs.

|

|
Fig. 13 - eggs, x 100, in
the uterus, inmature (up) half mature (below)
|
Fig. 14 - eggs, x
40. Those in the lower part of the picture are near the vulva
|

|

|

|
Fig 15 - start of the
spawning under the coverslip
|
Fig 16
- The seminal receptacle of one ovary, vulva and shed eggs
|
Fig 17 - Inverted colors
for a more clear view.
|
THE DEVELOPMENT OF
THE EGG IN THELASTOMATIDAE.
The
species of this family have a special ovular development. The
germinal cell begins to divide shortly after being shed, and
quickly reaches the state of
morula. That is to say the state of a compact spherical aggregation of
embryonic
cells.
The development of the embryo
then takes place within the egg generating a larva (L1) that develops
and moult
into a L2 larva, which produces a L3 larva as well. This embryonated
egg, with
an advanced larva must be eaten
by the future host (a nymph of
cockroach in our
case) in whose intestine the L3 moult to an L4 larva that gives rise
to
the adult.
This
implies, of course, that as is common in the parasitological diagnosis
in other animals or even in man that an examination of the ejections of
Periplaneta
can reveal, with no need of a dissection, which are the parasitized
individuals
and which are not. The following image
shows two eggs (easy to
identify
for those who have read this far), found in the examination of a drop
of
a water
suspension of Periplaneta
feces.

|
Fig. 18 - eggs in a drop
of a dilution in physiological solution of a fecal deposition from a
parasitized cockroach
Hammerschmidtiella,
left, and Thelastoma, right.
Obj. x 100 HI
|
THE
MALES FOUND IN THE SAMPLES

|
Dennis
van Waerebeke, a specialist in Thelastomoidea,
when describing a Leydinema of
Madagascar
reports the impossibility to surely assign a
certain
male to one certain species, because often the populations are mixed.
Exactly
the same happens with my samples of Durango. Except for a
male, probably assignable to Hammerschmidtiella,
because the population
of
females was compounded by only a few females of this genus, the other
two
specimens
that I found were in a mixed population. Anyway, one very
small male, probably an L4, is very similar to the male of Th. gipetiti van Waerebeke, 1987,
and can be confidently assigned to Thelastoma.
(Fig. 21)
There are at least 3 species of thelastoma assigned to cockroaches, but
only two to north American specimens. One is Th. bulloësi, the other is Th. periplaneticola
Leibesperger, 1960. Adamson, 1992 says that without the males,
the females of both species are not distinguishable from each
other. The only difference between the males is the lack of the
spicule in periplaneticola. My
example is too small to define this trait even with immersion
objective. In 1988
Adamson reports only 3 species for P.
americana; Th. bulloësi, L. appendiculata and H. diesingi. So I think that the
identifications applied here are good enough
There are
very few males; we only found one (fig.
19, at left)
in a population made exclusively of individuals of Hammerschmidtiella,
and
this is the reason why we think we can attribute it with some certainty
to
that genus.
It does
not have the pseudobulb so visible and
characteristic of the females, but a moderate widening in the base of
the
pharynx and before the isthmus. The pharynx is thus club shaped. The
isthmus is
longer that in the described female and the terminal bulb is less
important.
It is thus more similar to
the drawing of Chitwood.
Apparently
two testicles can be identified, aligned in a straight
line, one before the other, followed by a seminal vesicle that would
send the
spermatozoa to the cloaca. No alulae (cuticle features that aid in
embracing the
female) could be seen.
In this
image it is almost indistinguishable the only
chitinous spicule the Thelastomatidae males have. It is clearly seen in
another
somewhat different male found in another Periplaneta and which will be
illustrated next.
In order
to certify the species it is necessary to
locate and to describe certain perianal glands (named genital warts or
genital
papillae) that we have not been able to identify in our individual.
Anyway
this specimen is a good example of the general
structure of a male of the Thelastomatidae family.
An
important detail of the development of the Thelastomatidae is that
females are diploids (that is to say that they have a double gamete of
chromosomes,
originating from the fertilization of the haploid egg of the female
with the haploid
spermatozoon of the male), but all males are haploids (that is to say
that
they have
a single gamete of chromosomes). The diploid females can produce males
by
parthenogenesis,
without fecundation, which explains the haploidy of them. |
NOTE:
Any one that needs to refresh their knowledge around the
concepts of cellular nucleus, chromosomes,
mitosis, meiosis,
fertilization, parthenogenesis and ploidy can search Wikipedia,
entering
this link:
http://en.wikipedia.org/wiki/chromosome
and follow the numerous links that detail in very
accessible form all
concepts.
Using the finder at left of the page it is possible to reach the other
needed
terms.
I believe that this is a much
logical solution than to
establish a long
Glossary, with necessarily incomplete definitions.
Fig. 20. Unlike
the previous male whose abdomen finishes in conical form and
continued with one long tail similar to that of a female, in this
there is a
thinned end that forms one genital thickened papilla where a
characteristic spicule
is seen, and presents only a short tail although equally sharpened. The
thickness of both males is smaller than that of a female, and its
length is somewhat
shorter. Click the picture to see a labeled one.
This
other individual (apparently a male of Thelastoma by the characteristic
shape of its
pharynx) is evidently of very different structure, although it shares
important
characteristics with the previous ones.
Fig. 21. This
is one of two very small individuals found in a mixed population of
both genera. By its size it does not seem to be an adult form. Perhaps
it is a
larva in development. Cuticle was clearly ring-shaped. The mosaic was
composed
with pictures taken with the 100 x objective. Contrast and oblique
illumination
obtained with the Mathias Wedge. Click the image to see the labeled one.
MUTUALISTIC RELATIONSHIPS
The high incidence
(prevalence) of the infection by the protozoans Endamoeba and Nyctotherus,
and the nematodes Hammerschmidtiella, Thelastoma, and
even Leydinema
according to the data available, indicate that Periplaneta americana is not seriously affected
by their presence in the intestinal medium. It seems therefore that
this
is a clear
case of commensalism. All of them live in the shelter offered
by the cockroach intestine, and feed on the intestinal contents but
without harming their host.
Biological controls
of domestic cockroach invasions have been looked for, trying to
avoid the use
of chemical insecticides, which are really ecocides. It seems that the
most advantageous
proposal is the use of Steirneinema, another one genus of
nematodes, which
can really act as a control by killing the infected individuals.
Preparation of the nematodes
In my article in Micscape
of December, 2002, when discussing glycerin mounts I give the
suggestion to apply the method of Seinhorst,
the
standard method that almost all the professional nematodologists use.
It
seems also
to give a very good result, although it demands perhaps somewhat more
patience, by gathering the nematodes in 50 or 70%, alcohol and adding
10% glycerin. It is enough to let evaporate the liquid protected from
dust
so that when
the volume reduces to more or less a tenth, the animals are included in
an almost pure glycerin, in
which they can be finally mounted.
EPILOGUE
Those who
feel a vocation for these type of investigations can investigate other
insects,
dissecting other species of cockroaches, crickets, the so called mole crickets, of the Grillotalpydae family,
locusts and melolonthoid larvae of
many Coleoptera (called in
Spanish "gusanos blancos", in English "chafer grubs", or "
white grubs " and in French "vers blancs" or "larves de
Hanneton") which constitute a plague in gardens and orchards because
they
live buried, devouring the roots of the plants.
Other hosts are the
Coleoptera
of several families, their larvae, and centipedes (chilopoda) and
millipedes (diplopoda).
Even termites and ants host nematodes of the Thelastomatidae
family. It
is possible also to find nematodes of a near family: the
Rhigonematidae. But
its structure is very similar. The important differenc is that
the
Thelastomatidae have 8 buccal papillae and the Rhigonematidae, only 4.
To serve as a guide in the
generic determination of the individuals found in cockroaches the
following key
can be useful.
The key demands
an additional image to understand the difference
between Thelastoma and Leydinema. We took the following
one from the article of
van
Waerebecke on the Leydinema
genus. The original image is vertical, but
to save
space in screen we have placed it here in horizontal position.

|
|
Fig. 22 -
Intestine
of Leydinema portentosa van Waerebeke, 1987, showing the characteristic
intestinal loop and the "gastric cecum" distinctive of the genus.
|
The
Family Thelastomatidae is defined by the following characters:
OXYURIDA. They are inhabitants of the
digestive tract of insects
and some other arthropods, with 8 simple papillae in the external
circuit of buccal
papillae; one or no spicule in the males. The following genera have
been described
for the domestic cockroaches, specially Blatella germanica
and Periplaneta americana
KEY for the genera
1(2)
- Gastric cecum and intestinal loop..........................Leydinema
2(1)
– Without cecum nor loop.................................……………….3
3(4) -
One ovary, short and conical tail in both sexes
...........Blatticola
4(3)
- Two ovaries, tail long, thin and pointed…............……………5
5(6)
- Without pseudobulb...........………..………………Thelastoma
6(5)
- With pseudobulb and bulb.…..…….……Hammerschmidtiella
Blatticola inhabits
the small cockroach Blatella germanica, with incidence of 92% in
North
America.
The other 3 genera
can be found in Periplaneta orientalis. Hammerschmidtiella
has
apparently only 2 species in roaches. Thelastoma has 43
described
species, but
only 17 are accepted without problems, 23 need to be re-investigated
to define their situation, and 3 are badly described and it is not
known what their
true assignment is. Leydinema, has 3 recognized species (1998)
and
other 4
doubtful ones. D. van Waerebeke insists (1998) that the two really
important
traits that differentiate Leydinema
are the gastric cecum and the intestinal loop. In order to appreciate well
these characteristics, individuals must be observed preferably alive.
In wild cockroaches other genera have also been described, like Cephallobellus
Cobb, 1920; Coronostoma, Rao, 1958 and Severianoia,
(Schwenk,
1926) Travassos 1929. All the genera of this family parasitize also
other
insects and his larvae. An investigation of the Web using the generic
names
will allow data to be obtained on their morphology, hosts and
geographic
distribution.
The
next and very interesting step for the amateur microscopist
can be the investigation of the different parasites from frogs and
toads. As it
is difficult to obtain them in Cancún, the next article will
deal with a parasitic
species of
one nematode from the stomach of a small lizard
common in
the gardens of this city.
A NOTE
ON
Nyctotherus
As
a confirmation of what I said at the end of the second article in
this series (on the small significance of the number of individuals of Periplaneta
investigated until that moment), a pair of new dissections showed the
presence
in Cancún of a fauna totally similar to the one of Durango. In
the most
representative one I find a female of H. diesingi and two
females of T.
bulhoesi and in both cases a great amount of Nyctotherus. A
building
company has destroyed 16 of the 64 hectares of secondary forest in
front of
my
house. It is very probable that these invaders have a wild origin and
therefore
they have not been affected by the municipal fumigations of
insecticides, made
in the urbanized zone.
|




